项目文章 | 多组学探究肝癌微环境中影响免疫治疗好坏的肿瘤免疫屏障
2023-04-20 16:57:36, 欧易生物 上海欧易生物医学科技有限公司


2023年1月16日,中国科学技术大学附属第一医院刘连新团队在Journal of Hepatology(IF:30.083)发表了题为“Identification of a tumour immune barrier in the HCC microenvironment thatdetermines the efficacy of immunotherapy”的研究论文,该文章探究单细胞和空间转录组揭示肝癌微环境中影响免疫治疗好坏的肿瘤免疫屏障。

文章标题:Identification of a tumour immune barrier in the HCC microenvironment thatdetermines the efficacy of immunotherapy
单细胞转录组测序样本:6例肿瘤免疫治疗无反应患者中获得的肝癌肿瘤和邻近正常组织
空间转录组测序样本:8例接受抗PD-1治疗的肝癌患者的肿瘤切片(无应答者n=5;应答者n=3),相邻正常组织切片(n=3)
期刊:Journal of Hepatology
文章一作:刘尧
发表时间:2023年1月
方法:欧易生物10× Genomics scRNA-seq,10× Genomics Visium Spatial

只有少数HCC患者受益于ICB,这严重阻碍了免疫治疗在HCC患者中的应用。在这里,作者使用多组学方法鉴定了肿瘤免疫屏障(TIB)的空间结构,TIB是由肝癌微环境中的SPP1+巨噬细胞和CAFs相互作用形成的,这种结构限制了免疫细胞向肿瘤核心的浸润。体外实验中,低氧条件下巨噬细胞SPP1表达上调。此外,在SPP1+巨噬细胞特异性敲除小鼠肝癌移植模型、自发性肝癌小鼠和野生型小鼠肝癌移植模型中,阻断或敲除SPP1均能显著增强PD-1阻断效果,同时减少CAF浸润,增加T细胞的细胞毒性。总的来说,这些结果揭示了肿瘤微环境(TME)中TIB结构在免疫治疗疗效中的作用,并表明阻断SPP1+巨噬细胞与恶性肝细胞和CAFs的相互作用,联合抗PD-1治疗可以提高治疗疗效。

1、空间转录组揭示了与免疫治疗疗效相关的TIB微环境结构
为了探索免疫检查点封锁(ICB)治疗下的TME结构异质性,作者对8例接受抗PD-1治疗的HCC患者的肿瘤切片(无应答,n=5;应答者,n=3)和相邻正常组织切片(n=3)进行空间转录组测序。患者#1和#5的肿瘤组织与相邻的正常组织配对(图1A)。经过质量控制,根据无偏聚类和斑点特征,将相邻正常组织斑点分为HP、ADH1B高表达的肝细胞,RGS5、MYH11高表达的肌成纤维细胞/周细胞,PTPRC、CD3D、CD79A、COL1A1高表达的免疫/成纤维细胞,或FXYD2、KRT7高表达的胆管细胞。将肿瘤斑点分为肝细胞、免疫/成纤维细胞、肌成纤维细胞/周细胞、SPP1+巨噬细胞/CAFs(含SPP1、CD68、LUM和TIMP1)和恶性肝细胞(含AFP和APOA2)(图1B-G)。作者无法区分CAFs和SPP1+巨噬细胞,它们在同一点共定位多达10个细胞,这表明这两种细胞类型之间存在物理相互作用。有趣的是,围绕肿瘤细胞的SPP1+巨噬细胞/CAFs簇在ICB无反应者中产生了TIB结构(图1C),但在有反应者中没有(图1F)。SPP1+巨噬细胞/CAFs斑点在肿瘤中特异性富集,且ICB无反应者比例高于有反应者(图1H-I)。Bayspace增强空间数据显示,SPP1高表达区域在无应答者中高表达CD68和COLIAl,但在邻近正常组织或应答者中不表达(图1J)。此外,多重免疫组化(mIHC)染色显示,SPP1阳性细胞和αSMA阳性细胞在肿瘤周围非常接近,无应答者的双阳性细胞对数量明显高于应答者(图1K-L),这表明这两种细胞类型之间潜在的串音有助于形成与免疫治疗疗效相关的TIB结构。


图1 | 人肝细胞癌组织的空间转录组图谱揭示了ICB治疗的疗效
2、在ICB无反应的HCC患者中配对的相邻正常和肿瘤组织的单细胞转录组图谱
为了更好地了解ICB无反应患者中TIB的细胞组成,作者对从6例ICB无反应患者中获得的HCC肿瘤和邻近正常组织进行了scRNA-seq(图1A和2A)。在对scRNA-seq数据进行质量控制和双偶去除后,6例患者的83793个细胞用于后续分析(图2A)。作者通过Harmony整合所有来自肿瘤和邻近正常组织的细胞以纠正批处理效应,进行聚类分析并使用基于标记的注释来定义主要细胞类型,包括T/NK细胞、髓系细胞、B/浆细胞、成纤维细胞、内皮细胞和肝细胞/双能细胞(图2A-B)。在所有患者的肿瘤和邻近正常组织中均可发现六种主要细胞类型(图2C)。值得注意的是,髓系细胞和成纤维细胞的比例在肿瘤组织中显著增加(图2C)。组织、肿瘤和邻近正常组织之间浸润细胞类型的差异表明,TME的动态重塑在无反应的HCC患者中起着重要作用(图2D-G)。
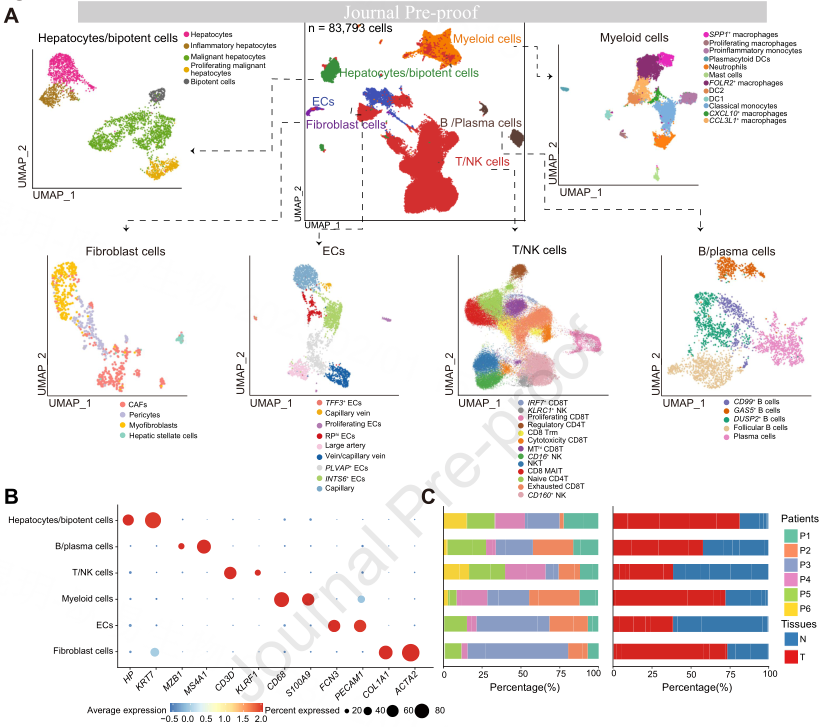

图2 | ICB无反应HCC患者中配对的相邻正常和肿瘤组织的单细胞转录组图谱
3、SPP1+巨噬细胞以TIB结构积聚在肿瘤组织中
为了进一步探索具有TIB结构的ICB无应答者骨髓细胞的异质性,作者应用类型特异性标记,将邻近正常组织中的3,034个细胞和肿瘤中的7,978个细胞分为12个亚型,包括5种表达CIQA和CD68的巨噬细胞亚型(SPP1+巨噬细胞、FOLR2+巨噬细胞、CXCL10+巨噬细胞、CCL3L1+巨噬细胞、增殖巨噬细胞),2种表达CD14的单核细胞亚型(经典单核细胞、促炎单核细胞),表达CSF3R的中性粒细胞,三种亚型的树突状细胞(树突状细胞;DC1, DC2,浆细胞样DC (pDCs)),以及表达TPSB2的肥大细胞(图3A)。为了证明作者对髓细胞的聚类是正确的,作者通过Harmony将作者的数据集与Sharma的[1]和Zhang的[2]数据集集成在一起。在Sharma的数据集中,带有SPP1的细胞主要在TAM2簇中,带有FOLR2的细胞主要在TAM1簇中。在簇Mp-C2-CIQA中表达SPP1和FOLR2,但在Mp-C2-CIQA簇中表达SPPI和FOLR2的细胞在Zhang的数据集中并没有重叠,这表明SPP1+巨噬细胞作为一个独立的亚型存在,并可能在TME中发挥重要作用。
然后,作者比较了肿瘤和邻近正常组织之间骨髓细胞亚型的比例(图3B-C)。肿瘤中SPP1+巨噬细胞的百分比显著增加(图3C),而相邻正常组织中pDC和DC1簇富集。正如之前报道的那样,肿瘤组织中FOLR2+巨噬细胞的比例往往高于邻近正常组织。
4、SPP1+巨噬细胞与肿瘤进展和低氧微环境相关
为了进一步研究大型队列中亚型改变的临床相关性,作者使用基于反卷曲的工具CIBERSORTx来预测在TCGA-LIHC队列中使用大量RNA-seq和其他来自基因表达综合(GEO)的独立队列中使用scRNA-seq量化的48种细胞类型的浸润。作者发现,在TCGA-LIHC数据集中,与邻近正常组织相比,肿瘤中SPP1+巨噬细胞的浸润明显丰富(图3D)。流式细胞术分析显示,无应答组肿瘤组织中SPP1+巨噬细胞浸润明显高于应答组(图3E)。此外,mIHC染色证实CD68共定位和SPP1蛋白表达的SPP1+巨噬细胞在HCC肿瘤中富集(图3F)。值得注意的是,在三个独立的队列中,SPP1+巨噬细胞浸润程度较高的HCC患者总生存期(OS)较短(图3G)。和较短的无进展生存期(PFS)在TCGA-LIHC队列(唯一可获得PFS信息的队列),而其他亚型巨噬细胞与患者预后无显著相关性。作者进一步证实,在HCC肿瘤芯片中,SPP1蛋白表达在肿瘤组织中特异性增加(因此在邻近的正常组织中不表达),并与预后不良有关(TMA;图3H-I)。此外,在TCGA-LIHC队列中,SPP1+巨噬细胞的比例在晚期增加(图3J),这表明SPP1+巨噬细胞可能促进肿瘤进展。
此外,SPP1+巨噬细胞具有不同于其他巨噬细胞亚型的功能。SPP1+巨噬细胞表达较高的转录因子(如EPAS1)与缺氧通路相关。FOLR2+高表达的巨噬细胞FOLR2、APOE、C2等;CXCL10+巨噬细胞高表达RNA编辑酶APOBEC3A,被认为可以促进促炎巨噬细胞的极化;CCL3L1+巨噬细胞高表达CD74、HLA-DR、HLADP和HLA-DQ,与抗原提呈相关。功能分析显示,糖酵解/糖异生、氧化磷酸化和HIF-1信号通路在SPP1+巨噬细胞中富集,而与吞噬体和抗原处理和呈递相关的基因在所有其他亚型的巨噬细胞中高度富集(图3K)。作者进一步发现体外缺氧可诱导SPP1在THP-1细胞上的高表达(图3L),而SPP1+巨噬细胞的吞噬评分最低,说明SPP1+巨噬细胞确实受缺氧影响,促进肿瘤进展,抑制吞噬。
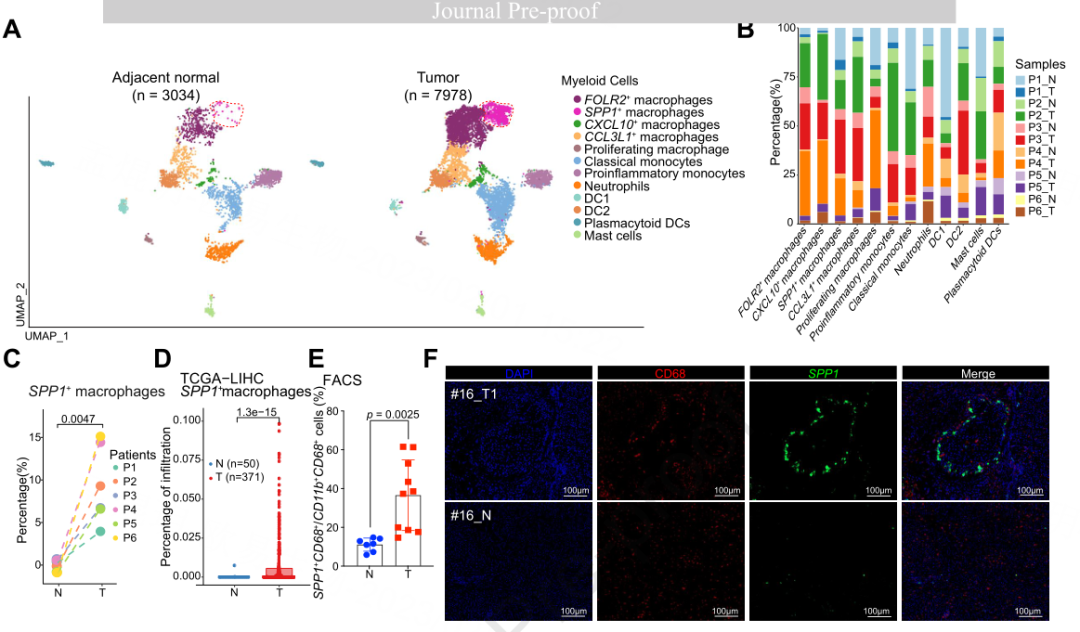

图3 | ICB无反应肿瘤和邻近正常组织中髓系细胞的特征
5、CAFs与SPP1+巨噬细胞的相互作用与TIB结构的形成有关
为了分析另一种TIB成分CAFs的细胞组成,作者在肿瘤和邻近正常组织的scRNA-seq数据集中将成纤维细胞亚聚为四种亚型(图4A),包括COLIA1、COLIA2、VCAN和TIMPI表达的CAFs;肌成纤维细胞表达SDO3、DSTN、MYH11和CD9;RGS5和NDUFA4L2表达的周细胞;肝星状细胞COLEC10, CD14,和HGF表达。然后,作者研究了相邻正常组织和肿瘤组织成纤维细胞亚型的变化,发现肿瘤组织中CAFs主要富集,而其他三种亚型成纤维细胞没有明显变化(图4B)。肿瘤组织中周细胞比例升高,但不明显,周细胞可促进肿瘤生长,并与内皮细胞共同影响肿瘤对免疫治疗的反应。CIBERSORTx反褶积分析显示,在TCGA-LIHC队列中,肿瘤样本中CAFs的浸润比例明显高于邻近正常组织(图4C)。在国际癌症基因组联盟(ICGC) HCC队列中,CAFs浸润较高的HCC患者OS较短(图4D),提示肿瘤组织中CAFs的富集可能与HCC患者预后不良有关。
然后,作者从scRNA-seq数据集中对具有CAF和SPP1+巨噬细胞特征的ICB无应答的每个空间聚类进行了标记,并突出了具有SPP1+巨噬细胞和CAF共定位的聚类(图4E, 4G)。SPP1+巨噬细胞/CAF空间聚类的特征评分也高于其他聚类(图4F、4H)。此外,SPP1+巨噬细胞和CAFs的签名评分在ST数据集(图4)和三个具有大量转录组的独立HCC队列中显示出显著的正相关。这些结果表明,SPP1+巨噬细胞和CAFs可能在TIB结构中发生物理相互作用。此外,作者发现在SPP1+巨噬细胞/CAF聚簇富集通路的位点上高表达的基因有助于TIB结构,这些基因富集于细胞外结构组织、细胞外基质组织以及细胞-细胞、细胞-基质和细胞-基质粘附(图4M)。在scRNA-seq数据集中,SPP1+巨噬细胞和CAFs中特异表达的基因的功能富集一致地揭示了一个共享的KEGG通路:ecm受体相互作用。这些结果表明,SPP1+巨噬细胞和CAFs的共定位可能与细胞迁移、粘附和ECM组织有关。
此外,作者用R包“NicheNet”评估了SPP1+巨噬细胞和CAFs之间假定的串扰,发现SPP1+巨噬细胞表现出较高的TGFB1、SPP1和IL1B配体活性。TGFB1在CAFs上与TGFBR1、TGFBR2、TGFBR3结合表达,SPP1在CAFs上与ITGA4、ITGA9、ITGAV、ITGB1、ITGB5相互作用,IL1B在CAFs上的配体为IL1R1。结果,靶基因胶原蛋白(COL1A1, COL1A2, COL3A1, COL4A1和COL5A1)、基质金属肽酶(TIMP1和MMP)和趋化因子(CCL3、4、5和CXCR4)在这些细胞中表达。这些靶点在结缔组织形成反应中起着重要作用,包括细胞外小生境纤维成分和基质金属蛋白酶。
值得注意的是,SPP1+巨噬细胞与CAFs配体-靶点相互作用的靶基因极有可能属于细胞因子-细胞因子受体相互作用、细胞外基质通路、TNF信号通路、IL-17信号通路(图4N)。然后,作者重点研究了前三个配体(TGFB1、SPP1和IL1B)和ECM相关靶点诱导的信号通路,发现了HIF1A和FOXO1等调控因子,这些调控因子富集在SPP1+巨噬细胞和CAFs的肿瘤组织中。促进ECM形成的靶基因,如TIMP1, LUM, FAP和PSOTN也在CAFs的肿瘤组织中富集(图4O)。
此外,从scRNA-seq数据中获得的SPP1+成纤维细胞前20个配体和CAFs前20个靶标的平均表达在SPP1+巨噬细胞/CAFs空间簇中高度富集(图4P-Q)。这些结果表明,SPP1+巨噬细胞和CAFs紧密位于TIB中,并通过这些配体受体相互作用促进TIB结构的形成。


图4 | 肿瘤和邻近正常组织中成纤维细胞的特征及其与SPP1+巨噬细胞的相互作用网络
6、TIB结构破坏恶性细胞和免疫细胞之间的相互作用,以限制对免疫治疗的反应
为了探索SPP1+巨噬细胞/CAFs在TIB中的潜在作用,作者评估了SPP1+巨噬细胞/CAFs和恶性肝细胞与其他空间簇的相互作用权重。作者发现,在ICB无应答者中,SPP1+巨噬细胞/CAFs与HCC的相互作用权重最强,但与免疫细胞的相互作用权重较弱,而在有应答者中,SPP1+巨噬细胞/CAFs、成纤维细胞和免疫细胞与HCC的相互作用权重相当(图5A-B)。根据已知免疫标记的分析,在ICB无应答者中,CD8+T淋巴细胞(CD3D和CD8A)和细胞毒性标记物(GZMB、GZMK和PRF1)对TIB结构包围的肿瘤(图5C)浸润有限或没有浸润(图5D),但在ICB应答者中,肿瘤中没有TIB结构和免疫细胞浸润(图5D)。
此外,SPP1+巨噬细胞/CAFs中髓系细胞和ECM特征的免疫抑制特征特征评分较高(图5E),提示SPP1+巨噬细胞可能与免疫抑制功能有关,CAFs可能与细胞外基质成分的生成有关。为了探索SPP1+巨噬细胞/CAFs在HCC免疫治疗中的功能作用,作者计算了T细胞炎症基因表达谱(GEP)在每个点的签名评分,以及每个空间簇中GEP基因的平均表达。作者发现TIB包膜的HCC组织中GEP评分较低,用于评估GEP评分的特征在TIB包膜的HCC组织中并不丰富(图5F-G)。此外,mIHC染色显示,SPP1+巨噬细胞倾向于定位在肿瘤边界,而CD8+ T细胞定位在TIB外,而不是在ICB无应答者的肿瘤核心(图5H),而SPP1+巨噬细胞浸润较少,CD8+T细胞浸润较多(图5I)。总之,这些结果表明,与SPP1+巨噬细胞相关的TIB结构的形成可能有助于HCC的免疫抑制微环境。


图5 | 在ICB无反应患者中,SPP1+巨噬细胞和CAFs的共定位作为肿瘤免疫屏障
7、靶向SPP1破坏TIB结构并使HCC对免疫治疗敏感
由于SPP1是一种分泌的糖磷蛋白,其表达与HCC的免疫抑制微环境密切相关,缺失SPP1可能会消除CAFs对免疫微环境的抑制作用,从而使杀伤T细胞浸润到肿瘤核心。为了验证这种可能性,作者将SPP1f/f和Lyz2-Cre小鼠杂交,生成了单核细胞中SPP1选择性缺失的小鼠系(SPP1f/f, Lyz2-Cre),然后使用SPP1f/f;Lyz2-Cre和SPP1f/f小鼠通过皮下移植Hepa 1-6细胞建立了一个同基因肝肿瘤模型。与SPP1f/f小鼠相比,抗PD -1处理的SPP1f/f, Lyz2-Cre小鼠的肿瘤生长显著减少,治疗效果更好(图6A)。流式细胞术分析显示,在SPP1f/f的肿瘤区,抗PD -1治疗显著增加肿瘤浸润CD8+ T细胞(图6B)和颗粒酶B+ CD8+ T细胞(图6C)的数量;Lyz2-Cre老鼠。mIHC染色结果显示,颗粒酶B对SPP1肿瘤区CD8+ T细胞的染色强度;PD-1处理的Lyz2-Cre小鼠高于其他各组(图6D-E)。
此外,作者还研究了抗SPP1和抗PD1联合治疗在Hepa 1-6肝肿瘤模型中的作用。与SPP1条件敲除小鼠的结果相似,在具有Hepa 16肿瘤的免疫功能小鼠中,联合抗SPP1和抗PD -1治疗可诱导肿瘤生长减缓,与单独使用每种治疗相比,反应率显著提高(图6F)。联合抗SPP1和抗PD -1治疗也显著增加了肿瘤区域颗粒酶B+和CD8+T细胞的数量(图6G-H)。重要的是,作者发现SPP1条件敲除和抗SPP1治疗组aSMA+成纤维细胞表达降低。接下来,作者在同基因免疫能力小鼠中生成H11LNL-Myc, Alb-Cre基因工程小鼠模型(GEMM)衍生的原位肝癌模型(GDL),然后用抗SPP1和/或抗PD -1治疗荷瘤GDL小鼠。抗SPP1或抗PD -1治疗可略微提高总生存率(图6I)。值得注意的是,抗SPP1和抗PD -1组合显著提高了荷瘤GDL小鼠的总体生存率(图6I)。此外,联合抗SPP1和抗PD -1治疗也显著降低了aSMA+成纤维细胞的比例,并增加了肿瘤区域CD8+T细胞的比例(图6J)。此外,作者验证了SPP1的阻断或条件敲除小鼠巨噬细胞特异性SPP1缺失。从Trp53KO/C-MycOE肿瘤模型中获得的肿瘤细胞对抗PD -1治疗不敏感,结果一致。这些结果表明,阻断SPP1可以增强抗PD -1免疫治疗的疗效。
因此,SPP1可能作为一种有价值的生物标志物,用于预测需要免疫治疗的HCC患者的结局,并且靶向SPP1进一步使HCC对免疫检查点抑制剂敏感。


图6 | TIB对SPP1+巨噬细胞的破坏促进了抗PD -1治疗小鼠模型的疗效

1. 整合多组学确定了肝癌微环境中肿瘤免疫屏障(TIB)的空间结构,有助于免疫治疗的疗效。
2. SPP1+巨噬细胞与癌相关成纤维细胞(CAFs)的相互作用促进了TIB结构的形成,限制了免疫细胞浸润到恶性区域。
3. 阻断小鼠SPP1或巨噬细胞特异性缺失SPP1可破坏TIB结构,使HCC对免疫治疗敏感。
Reprogramming of Endothelial Cells Drives Immunosuppressive Macrophages in
Hepatocellular Carcinoma. Cell 2020;183:377-394.e21.
Immune Cells in Hepatocellular Carcinoma. Cell 2019;179:829-845.e20.
猜你想看
1、项目文章 | 单细胞测序鉴定猪流行性腹泻病毒感染的猪小肠的细胞类型和转录组景观
2、项目文章 | 同济医院张子强团队运用AFADESI空间代谢组与微生物组联用探究肠道菌抗癌新机制~

END
排版人:小久
原创声明:本文由欧易生物(OEBIOTECH)学术团队报道,本文著作权归文章作者所有。欢迎个人转发及分享,未经作者的允许禁止转载。


04-22
CCMT2026开展即高能 | Equator-X™ 双模式测量仪引爆全场04-21
告别预测偏差!Percepta自建专属训练库,pKa预测更准更快04-21 ACDLabs 李丹
世界地球日,查看地球的【愿望清单】04-21
【前沿激荡,智汇北京】IGC 2026圆满落幕,益世科生物共绘细胞基因治疗新蓝图04-21
会议预告|华大吉比爱邀您共赴第六届北京临床质谱论坛04-21 华大吉比爱
叮咚!您有一份来自地球的“绿色盲盒”待拆封04-21 安捷伦科技
聚焦小核酸与多肽前沿分析:第一届创新药物研发与质量评价技术交流会在沪圆满举办04-21 安捷伦科技
糖基化全流程质控:消耗品方案一站式解决04-21 安捷伦科技
药物分析排忧解难系列 | 自动柱前衍生化方案,完美应对样品复杂前处理04-21 安捷伦科技
“衣”路前行,“安”全守护 | 安捷伦 PFAS 解决方案守护运动健康04-21 安捷伦科技
零压力,低成本 | 安捷伦官方认证翻新仪器开启“以租代售”新模式04-21 安捷伦科技
ESCMID 2026 爆点 | MiSeq i100系列携6项病原精准测序应用数据亮相04-21
高效+精准,VA-5000 让实验室气体分析从此不同!04-21 HORIBA
突破制药检测瓶颈,欧美克NS-180Z系列纳米粒度及电位分析仪重磅登场04-21 欧美克仪器
NS-180Z Pro丨纳米粒度电位分析仪04-21 欧美克仪器
【每天学点粒度知识】动态光散射测量粒径应用案例04-21
还有人不知道吗?代谢组才是多组学联用的灵魂04-20
【星品周 | 好礼翻倍】外泌体富集及miRNA提取攻略04-20
烷基汞检测技术及应用网络研讨会04-20 普立泰科