项目文章 | GC-MS代谢组学助力南京农大大豆疫霉根腐病防御机制研究
2021-02-04 11:44:27, 多层组学定制服务 上海欧易生物医学科技有限公司

点击上方“蓝色字体”关注我们

前言
欧易/鹿明生物合作客户南京农业大学作物遗传与种质创新国家重点实验室邢邯课题组在Frontiers in Plant Science发表题为“Metabolomics Analysis of Soybean Hypocotyls in Response to Phytophthora sojae Infection”的研究成果,通过GC-MS非靶向代谢组学、转录组学联合分析揭示大豆对P.sojae代谢水平上的防御反应,这项工作强调了植物-病原相互作用代谢组学研究的价值,并为大豆-P.sojae互作的关键代谢因素的研究铺平了道路。

中文标题:大豆下胚轴对大豆疫霉侵染反应的代谢分析
研究对象:大豆下胚轴
发表期刊:Frontiers in Plant Science
影响因子:4.402
合作单位:南京农业大学
运用欧易/鹿明生物技术:GC-MS非靶向代谢组学(由鹿明生物提供技术支持)
研究背景
大豆是世界上最重要的经济作物和油料作物之一。由大豆疫霉引起的大豆疫霉根腐病(P.sojae)是世界上大多数大豆产区的主要病害。目前,代谢组学尚未应用于大豆—P.sojae 相互作用以及Rps(抗P.sojae 基因)介导的防御反应机制的研究中。因此在代谢水平上大豆对大豆疫霉根腐病的防御反应机制鲜为人知。
本文利用代谢组学研究了2个大豆品系(1个为含有RpsJS 基因的抗性大豆系,1个为敏感大豆系)在被大豆疫霉根腐病(P.sojae)感染后2个时间点下胚轴代谢物的变化。并将代谢组学数据与转录组学数据(由RNA-Seq获得)整合,从分子水平上了解代谢水平的防御反应机制,发现新的抗病代谢产物及其相应的调控基因,为今后研究RpsJS 介导的防御反应机制以及大豆抗病育种提供更多的选择和思路。
研究思路

研究结果
1.大豆下胚轴对P.sojae侵染反应的代谢组学数据综述
利用GC-MS非靶向代谢组学技术,共鉴定出311个代谢物。结果表明,每组的生物学重复的样本都聚集在一个小区域(图1A),表明重复性较高。R品系的所有样品均聚集在图的左上角,S品系的所有样品均聚集在图的右下角,表明R品系和S品系的代谢物谱有显著差异。
相对于S品系中代谢物的表达情况,50种代谢物的表达水平在R品系的两个时间点都有显著差异累积(图1B)。图1C所示为各对照组中VIP值排名前5的差异代谢物,这些化合物,特别是糖(甜酒糖酶、异麦芽糖、山梨糖)和次生代谢产物(吲哚-3-乙酰胺、次黄嘌呤),可能在大豆对P.sojae侵染反应中发挥重要作用。图1D的韦恩图说明,不同组既有不同的差异累积代谢物(DAMs)也有共享的DAMs。

图1 | 差异代谢物的相关统计分析
(A)大豆疫霉下胚轴代谢产物的主成分分析,每组8次生物学重复
(B)显著差异累积代谢物(DAM)的数量。黄色(上调)和蓝色(下调)
(C)各比较组TOP5 VIP值的DAMs一览表
(D)DAMs的韦恩图分析
左框为下胚轴感染样品和相应的对照组之间的DAMs水平,右框为R品系和S品系之前的DAMs水平。R12:R-12h-IN vs.R-12h-CK;R36:R-36h-IN vs.R-36h-CK;S12:S-12h-IN vs.S-12h-CK;S36:S-36h-IN vs.S-36h-CK;RS12:R-12h-CK vs.S-12h-CK;RS36:R-36h-CK vs.S-36h-CK。IN:接种P.sojae ,CK:对照组
通过韦恩图和热图的结合有助于推测每个代谢物在防御反应中的作用(图2)。其中28个代谢物不仅在R品系和S品系之间有差异积累,在P.sojae侵染前后也有差异积累(图2中的红色字体)。

图2 | 从韦恩图的每个子集数据得到的DAMs热图
红色代谢物表示P.sojae 侵染后R品系和S品系之间差异累积代谢物
黄色(上调)和蓝色(下调),每个水平行代表一个DAMs名称
R12:R-12h-IN vs.R-12h-CK;R36:R-36h-IN vs.R-36h-CK;S12:S-12h-IN vs.S-12h-CK;S36:S-36h-IN vs.S-36h-CK;RS12:R-12h-CK vs.S-12h-CK;RS36:R-36h-CK vs.S-36h-CK。IN:接种P.sojae ,CK:对照组。
2.P.sojae侵染对大豆下胚轴碳水化合物代谢的影响
21种代谢物参与碳水化合物代谢途径,如淀粉和蔗糖代谢、乙醛酸和二元酸代谢、TCA循环等。
基于KEGG通路,联合转录组学数据和代谢组学数据进行综合分析。调节海藻糖合成的TPP(海藻糖6-磷酸酶,Glyma13g01910和glyma04g11250)基因在S36、R12和R36中的表达发生了变化,与海藻糖的变化相一致。R36中蔗糖的下调可能是由于R36中编码蔗糖裂解酶的基因表达上调,如AGLU(Glyma09g03250)和INV(β-呋喃糖苷酶/转化酶、Glyma15g02850、Glyma17g34570和Glyma17g34590)。但其下游产品(D-果糖)含量的变化尚未确定(图3A)。
S品系在被P.sojae侵染36 hpi时,琥珀酸、L-苹果酸和柠檬酸等有机物水平以及与TCA循环相关的柠檬酸合成酶(CS)、苹果酸脱氢酶(MDH)、异柠檬酸裂解酶(ICL)和乌头酸水合酶(ACO)基因对大豆疫霉侵染有响应。但调节草酸和2-甲基富马酸代谢的基因表达没有变化(图3B)。

图3 | 碳水化合物代谢途径图及其相关的DEGs(差异表达基因)和DAMs
(A)淀粉和蔗糖代谢途径
(B)TCA循环、乙醛酸盐和二元酸盐代谢途径
DAMs是黑色粗体字体,无显著差异的累积代谢物为黑色字体,未识别的代谢物为灰色字体,基因名缩写为斜体。红色表示上调DEGs,绿色表示下调DEGs,黄色表示上调DAMs,蓝色表示下调DAMs。矩形代表R12、R36、S12、S36、RS12和RS36。R12:R-12h-IN vs.R-12h-CK;R36:R-36h-IN vs.R-36h-CK;S12:S-12h-IN vs.S-12h-CK;S36:S-36h-IN vs.S-36h-CK;RS12:R-12h-CK vs.S-12h-CK;RS36:R-36h-CK vs.S-36h-CK。IN:接种P.sojae ,CK:对照组。
3.P.sojae侵染对大豆下胚轴氨基酸代谢的影响
联合分析转录组数据和代谢组数据,代谢物含量的变化与编码相应催化酶的基因表达的变化之间的相关性不高(图4,5)。然而,研究确定了一些基因,包括PAOs(多胺氧化酶)和SPDEs(精脒合酶),参与腐胺和亚精胺之间的转换;ARGI(精氨酸酶)参与精氨酸代谢(图4);
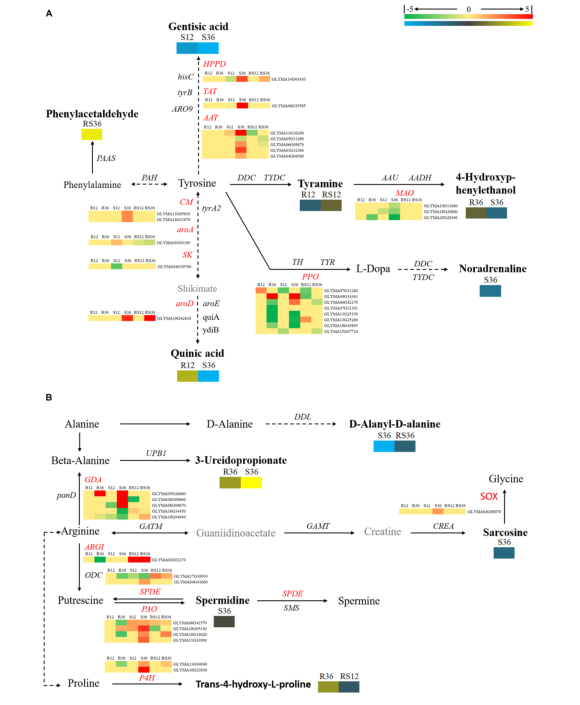
图4 | 氨基酸代谢途径图及其相关的DEGs和DAMs
(A)酪氨酸和苯丙氨酸代谢途径
(B)β-丙氨酸、脯氨酸和精氨酸代谢途径
DAMs是黑色粗体字体,无显著差异的累积代谢物为黑色字体,未识别的代谢物为灰色字体,基因名缩写为斜体。红色表示上调DEGs,绿色表示下调DEGs,黄色表示上调DAMs,蓝色表示下调DAMs。矩形代表R12、R36、S12、S36、RS12和RS36。R12:R-12h-IN vs.R-12h-CK;R36:R-36h-IN vs.R-36h-CK;S12:S-12h-IN vs.S-12h-CK;S36:S-36h-IN vs.S-36h-CK;RS12:R-12h-CK vs.S-12h-CK;RS36:R-36h-CK vs.S-36h-CK。IN:接种P.sojae ,CK:对照组。
ALDHs(醛脱氢酶),参与吲哚乙酸下游代谢;HMT(同型半胱氨酸S-甲基转移酶)和METE(5-乙基四氢叶酸同型半胱氨酸甲基转移酶)参与N-甲酰-L-蛋氨酸上游代谢,可能在大豆对大豆赤霉病的反应中发挥重要作用(图5)。

图5 | 氨基酸代谢途径简图及其相关的DEGs和DAMs
(A)色氨酸代谢途径
(B)半胱氨酸和赖氨酸代谢途径
DAMs是黑色粗体字体,无显著差异的累积代谢物为黑色字体,未识别的代谢物为灰色字体,基因名缩写为斜体。红色表示上调DEGs,绿色表示下调DEGs,黄色表示上调DAMs,蓝色表示下调DAMs。矩形代表R12、R36、S12、S36、RS12和RS36。R12:R-12h-IN vs.R-12h-CK;R36:R-36h-IN vs.R-36h-CK;S12:S-12h-IN vs.S-12h-CK;S36:S-36h-IN vs.S-36h-CK;RS12:R-12h-CK vs.S-12h-CK;RS36:R-36h-CK vs.S-36h-CK。IN:接种P.sojae ,CK:对照组。
4.P.sojae侵染对大豆下胚轴次生代谢物的影响
变化的次生代谢途径主要集中在以苯甲酸降解(图6A)、嘌呤代谢(图6B)和类黄酮、异黄酮生物合成(图7)。其中,只有分别参与4-羟基苯甲酸和4-羟基苯甲醛上游代谢的4CL(4-库马酸-CoA连接酶)和CYP71D9(细胞色素P45071D9)在S品系中显示出差异表达(图6A)。XDHs(黄嘌呤脱氢酶/氧化酶,Glyma13g41520和Glyma15g03870)参与了次黄嘌呤下游代谢,在R36和S36中上调。其余参与嘌呤代谢的差异表达基因仅对S36中的大豆疫霉感染有反应(图6B)。

图6 | 次级代谢途径图及其相关的DEGs和DAMs
(A)以苯甲酸降解为中心的代谢途径
(B)嘌呤代谢途径代谢途径
DAMs是黑色粗体字体,无显著差异的累积代谢物为黑色字体,未识别的代谢物为灰色字体,基因名缩写为斜体。红色表示上调DEGs,绿色表示下调DEGs,黄色表示上调DAMs,蓝色表示下调DAMs。矩形代表R12、R36、S12、S36、RS12和RS36。R12:R-12h-IN vs.R-12h-CK;R36:R-36h-IN vs.R-36h-CK;S12:S-12h-IN vs.S-12h-CK;S36:S-36h-IN vs.S-36h-CK;RS12:R-12h-CK vs.S-12h-CK;RS36:R-36h-CK vs.S-36h-CK。IN:接种P.sojae ,CK:对照组。
并且,似乎参与苯丙酸生物合成途径的关键基因,如C4H(香豆酸 4-羟化酶)、PALs(苯丙氨酸氨解酶)和4CLs(4-香豆酸-CoA连接酶)对R品系中的大豆疫杆菌感染没有反应(图7)。

图7 | 异黄酮生物合成一个分支代谢途径图及其相关的DEGs和DAMs
DAMs是黑色粗体字体,无显著差异的累积代谢物为黑色字体,未识别的代谢物为灰色字体,基因名缩写为斜体。红色表示上调DEGs,绿色表示下调DEGs,黄色表示上调DAMs,蓝色表示下调DAMs。矩形代表R12、R36、S12、S36、RS12和RS36。R12:R-12h-IN vs.R-12h-CK;R36:R-36h-IN vs.R-36h-CK;S12:S-12h-IN vs.S-12h-CK;S36:S-36h-IN vs.S-36h-CK;RS12:R-12h-CK vs.S-12h-CK;RS36:R-36h-CK vs.S-36h-CK。IN:接种P.sojae ,CK:对照组。
研究结论
本研究通过GC-MS非靶向代谢组学检测大豆在一段时间过程中的代谢变化,为大豆对大豆疫霉病的防御反应提供了新的视角。代谢物包括糖(左旋葡聚糖,红糖,海藻糖,异麦芽糖),有机酸(草酸,孜然酸和2-甲基富马酸),氨基酸衍生物(酪胺,糖阿片,N-甲酰-L-蛋氨酸,N-α-乙酰-L-鸟氨酸,苯乙醛吲哚-3-乙酰胺,4-羟基苯甲酸,反式-4-羟基-L-脯氨酸和S-羧甲基半胱氨酸)和次级代谢产物(大豆苷元、次黄嘌呤和辛醛)都可能参与防御反应。
尽管本次研究中的转录组和代谢组数据之间的联合和相互依赖性较差,但仍然发现了一些有价值的差异表达的基因,如INV、CHS、IFS,XDH等,它们参与了潜在抗性物质的调控。实验中的这些发现为后续的研究项目指明了方向,例如RpsJS介导的防御机制的研究。此外,这些发现的进一步研究将促进大豆抗大豆疫霉病品种的选育。
小鹿推荐
本文研究了南农10-1(抗性系,R)和06-070583(敏感系,S)两个大豆品系在P.sojae处理后2个时间点(12和36hpi)下胚轴代谢谱的变化以及两个品系之间的差异代谢物。50种代谢物的水平在R系和S系之间存在差异,且在P.sojae处理后鉴定出90种差异累积代谢物(DAMs)。共有28个DAMs,它们不仅在R系和S系之间有差异积累,而且在P.sojae处理后也有差异积累。基于不同品系在不同时间点在被P.sojae侵染后代谢物含量的变化,研究者推测这些代谢物包括糖(单糖和低聚糖)、有机酸(草酸、孜然酸)、氨基酸衍生物和其他次生代谢物(甘露醇、辛醛、丙烯酰胺、次黄嘌呤、大豆黄酮等)可能参与大豆对P.sojae代谢水平上的防御反应。在本研究中,对转录组学(由RNA-Seq获得)和代谢组学数据的联合分析表明,大豆对P.sojae的防御响应,在代谢水平和转录水平之间没有较好的关联和互相依赖性。这项工作强调了植物-病原相互作用代谢组学研究的价值,并为大豆-P.sojae互作的关键代谢因素的研究铺平了道路。
文末看点
上海鹿明生物科技有限公司,一直专注于生命科学和生命技术领域,是国内早期开展以蛋白组和代谢组为基础的多层组学整合实验与分析团队。本篇是针对大豆的防御反应研究,同时在GC-MS非靶向代谢组学研究方向还有以下的方向:
农药对水稻微生物群与氮循环影响
 项目文章PBP | 浙江工业大学傅正伟运用GC-MS非靶向代谢组学和16S探究水稻根际微生物群落影响
项目文章PBP | 浙江工业大学傅正伟运用GC-MS非靶向代谢组学和16S探究水稻根际微生物群落影响
AgNPs暴露对土壤微生物影响
项目文章EST | GC-MS代谢组学助力南京大学赵丽娟课题组再登环境科学TOP期刊
白灵菇菌丝体生理成熟潜在指标研究
客户文章 |中国农科院胡清秀博士课题组运用GC-MS代谢组学对白灵菇菌丝体生理成熟过程中的代谢轮廓谱研究
☆欢迎长按扫码添加鹿明生物技术工程师微信,深入探讨GC-MS非靶向代谢组学研究

// 长按扫码即可咨询 //
获取本篇文章原文
咨询鹿明生物GC-MS代谢组学工程师
实验样本准备事项
参考文献:
Longming Zhu, Yang Zhou, et al. Metabolomics Analysis of Soybean Hypocotyls in Response to Phytophthora sojae Infection.[J]. Frontiers in plant science, 2018,9:1530.
猜你还想看
END
欢迎转发到朋友圈
本文系鹿明生物原创
转载请注明本文转自鹿明生物

我知道你在看哟


点“阅读原文”查看更多精彩
04-16 德国元素
走进信阳师范大学04-16
应用分享丨Micro-XRF技术在高性能材料研发中的革新应用04-16
蓄势聚力 蓬勃葳蕤 |《皖仪季风·春生》四月刊04-15 WAYEAL皖仪
血液专题 | 血液全场景解决方案上线!直播教学帮您深入了解这些技术怎么用~04-15
凝聚科技力量·共筑交通安全:欧美大地引领交通安全设施测试创新发展04-15 公路部
创新2024 | 无氯仿提取RNA,让科研更安心!04-15 赛默飞生命科学
创新2024 | 细节决定成败:RNA实验中RNase防控要点全解析04-15 赛默飞生命科学
创新2024 | 告别低效转染,Neon NxT电转染系统为科研创新加速!04-15 赛默飞生命科学
探秘创新之旅,赛默飞“灵感”系列笔记本助力发展新质生产力,好礼等你来拿!04-15 赛默飞生命科学
创新2024 | 革命性突破!Bigfoot流式细胞分选仪:单细胞组学研究的创新引擎04-15 赛默飞生命科学
【新品上市】1.8μm BioCore Glycan糖型分析色谱柱04-15
塑料实验室光源暴露试验荧光紫外灯方法04-15
浙江万马高分子材料集团有限公司对我炭黑含量测试仪认可04-15
感谢安徽工程大学对我司同步热分析仪的认可04-15
聚碳酸酯PC再生塑料性状及性能要求04-15
带通滤光片产品介绍04-15 韵翔光电
奖品更多?名额增加?色谱能力验证大赛第二届来袭!04-15 市场宣传部
【展会预告】第25届中国环博会04-15 元析仪器
元析仪器 | 2024德国慕尼黑分析生化博览会圆满落幕04-15 METASH