Cell Metabolism重磅综述 | 肠-肝轴的病理生理学概念和临床意义(上)
2022-12-09 00:23:36, 麦特绘谱 麦特绘谱生物科技(上海)有限公司
过去几年中,肠道和肝脏之间的双向交互作用已经成为实验性肝病的关键影响因素,越来越多的临床证据表明,在肝病和肝病以外的疾病中都出现了肠-肝轴的紊乱。来自奥地利因斯布鲁克医科大学的Herbert Tilg教授团队,讨论了肠道和肝脏之间双向交互的实验和临床结果,综述了健康和肝病中的特定介质变化;阐述了肠道和肝脏在健康和疾病中复杂的代谢相互作用,对未来靶向治疗肝脏疾病提供理论基础,该综述发表于Cell Metabolism 。

肠道屏障受损是肠-肝轴改变的先决条件
肠道屏障保护身体免受潜在的有毒代谢物、细菌及其抗原的影响。因此,对肠道屏障和共生菌群的免疫控制至关重要。多种因素促进了这种多层面和动态的宿主-菌群共生关系。例如,菌群间的互作决定了成熟的肠道菌群的发展,免疫反应必须由宿主的细胞和体液免疫系统很好地平衡。近年来,饮食成为哺乳动物肠道屏障的一个关键调控因素(图1)。
图1. 肠道屏障:肠-肝轴的守门员
饮食与肠道屏障
受损的肠道屏障反映了肠-肝轴的“第一击”,这可能是由饮食应激和相关的菌群失调引起的。过去几年,饮食,特别是高脂肪饮食(HFD)已成为损害健康志愿者肠道屏障和内毒素血症的最重要的诱因。与这一概念相一致的是非洲人生活方式的城市化(即从农村向城市生活方式和饮食的转变)引发了机体对炎症的易感性。由内毒素等细菌成分引起的代谢性内毒素血症已被认为是一个将肠道屏障损伤与全身炎症联系起来的疾病。HFD还利用乳糜微粒通过肠道屏障运输内毒素。此外,摄入过多果糖会导致十二指肠道屏障功能受损,并可导致内毒素血症和非酒精性脂肪性肝病(NAFLD)。西方饮食中富含的膳食乳化剂也会破坏肠道屏障,并在小鼠中引发轻度全身炎症和肠道菌群失调。还发现,宿主的调控机制也会影响肠道屏障,例如在T2D(2型糖尿病))和NAFLD中,血糖浓度调控着肠道屏障功能障碍。
菌群与肠道屏障
与现在的粪便相比,1000 - 2000年前的古粪便表现出明显的菌群基因集富集。因此,肠道菌群被认为是控制肠道屏障和代谢性疾病的关键因子。例如,抗生素处理和在ob/ob小鼠中敲除CD14(一种Tolllike受体4 [TLR4]微生物感应的辅助受体)可以消除代谢功能障碍和全身炎症。共生菌通过多种机制有助于维持健康的肠道屏障,如粘液的维持和抗菌肽及短链脂肪酸(SCFAs)的产生。厚壁菌和拟杆菌被认为发挥有益的作用,但变形菌在粘膜表面却具有炎症作用,破坏肠道屏障。例如,补充普氏栖粪杆菌Fecalibacterium prausnitzii和嗜黏蛋白阿克曼菌Akkermansia muciniphilia(AKK菌)会影响小鼠的肠道菌群,并修复肠道屏障。AKK菌可增加肥胖小鼠和人类的粘液厚度,增加紧密链接表达,并改善代谢功能。同时粘膜真菌通过从小鼠的CD4+T细胞中产生白细胞介素22 (IL-22)来加强肠道屏障。
免疫介质与肠道屏障
20多年来,细胞因子已经被证实可以控制肠道屏障功能。例如,TNF、IFNg和IL-1b、内毒素、趋化因子成为损害肠道屏障的关键介质。反过来,IL-22可维持肠道稳态和肠道屏障功能;IL-18、IL-21等也有类似报道。维持肠道屏障的免疫细胞主要是特异性自我维持的巨噬细胞,它们紧密定位于血管,这种特化巨噬细胞的成熟高度依赖于肠道菌群,也强调了肠道屏障的维持需要肠道菌群的相互作用。
肠道-血管屏障
肠道-血管屏障(GVB)可防止细菌从肠道传播。GVB主要由内皮细胞组成,周围是周细胞和肠道胶质细胞,受损的GVB伴随着质膜膜泡关联蛋白(PV1)水平的升高。β-catenin(β-连环蛋白)信号通路控制血管通透性,β-catenin的的过量表达可保护小鼠免受沙门氏菌感染。在高脂诱导的NAFLD模型中,GVB受损,随后细菌转移至肝脏。HFD诱导的菌群失调导致了GVB的破坏,因为将HFD喂养的小鼠粪便转移至对照喂养的小鼠后,发现其也会破坏GVB。还发现,乙醇喂养小鼠可使PV1表达增加,而口服AKK菌可降低PV1的表达。然而,β-catenin的过表达并不能预防乙醇诱导的肝损伤,这表明GVB的特定功能。总之,肠道屏障包含复杂的生理和功能结构,从完整的粘液层到上皮紧密连接、宿主-菌群互作和GVB,它们共同维持健康,并影响多种肝脏疾病(图1)。
饮食信号干扰肠-肝轴稳态
饮食中的蛋白质和碳水化合物在肠道中被宿主及其共生菌群加工,然后通过门静脉进入肝脏。尽管这些产物或代谢物中大多数是生命所必需的,例如能量代谢,但许多产物或代谢物有可能引发或恶化炎症性疾病,特别是在过量摄入时。接下来主要探讨在肠-肝轴中起作用并导致肝脏疾病的促炎和抗炎饮食成分/代谢物。更重要的是,饮食对这种肠-肝轴的影响很少能脱离菌群而单独存在。
饮食是肠道菌群的主要调节器
营养和膳食成分是调节胃肠道菌群结构和功能的关键因素之一。早期的一项研究表明,与典型西方饮食的意大利儿童相比,非洲儿童表现出普雷沃氏菌丰度增加,而潜在致病菌厚壁菌和肠杆菌的数量减少。同样,HFD对肠道菌群的组成也产生了显著影响。另外的几项研究表明,饮食干预也会在短期内影响肠道菌群。通常存在于加工食品中的膳食乳化剂不仅会改变肠道屏障,还会严重改变肠道菌群,减少多样性和SCFAs。使用诸如植物纤维等健康食品的膳食干预,则对肠道菌群和相关血浆蛋白质产生有益影响。目前,大量的临床研究已经证明了饮食和肠道菌群之间的重要相互作用。例如,来自美国和土耳其的晚期肝病患者中,富含咖啡、谷物、茶、蔬菜和发酵乳品的饮食与较高的微生物多样性有关,饮食还会影响住院风险等。
生物节律控制肠道菌群共生和能量代谢
生物节律因素对宿主生理和代谢产生关键的影响。在过去的几年里,肠道菌群在过去几年被认为是传递昼夜节律饮食信号的一个重要因素。例如,膳食成分,特别是菌群代谢物,如SCFAs和BAs(胆汁酸),可以调控肠道菌群和昼夜节律通路之间的联系,而西方饮食则会扰乱小鼠肠道菌群的昼夜节律。膳食种类和节律性影响小鼠的肠道菌群和上皮转录反应。还发现,饮食诱导的肥胖会损害回肠菌群组成和转录组的昼夜变化,而时间限制的喂养可以改善这种变化。但是目前大多数人类研究没有考虑到昼夜节律的影响,未来的研究需要注意这个方面。
促炎的饮食
饮食中的饱和长链脂肪酸进入上皮来源的乳糜微粒中,通过淋巴系统吸收(由肠道菌群促进),经由血液进入肝脏。这些饱和脂肪酸是强效促炎营养物质,通过炎症小体信号通路激活炎症级联反应。HFD处理导致小鼠肠内产生IL-17的T细胞增加,同时调节型T细胞减少,而抗炎饮食则恢复了肠通透性和内毒素血症。西方饮食通常富含多不饱和脂肪酸,如n-6多不饱和脂肪酸,这样的饮食则引发老年小鼠的免疫细胞积累和肠道失调。喂食14个月以上的高脂/高胆固醇(HFHC)饮食导致小鼠的肝脏脂肪变性、肝脏炎症、纤维化和肝细胞癌(HCC),且将饮食诱导的肠道菌群失调转移到无菌小鼠中重现这种表型。因此,NAFLD中肝脏脂肪变性和肝脏炎症的演变是饮食诱导炎症的典型路径,表明营养物质过剩会沿肠-肝轴传播疾病。
除了脂类外,单糖也会导致肠-肝轴的病理。例如,果糖过量导致新生脂肪生成、非酒精性脂肪性肝炎(NASH)和HCC,这与肠道屏障受损、内毒素血症和激活MyD88介导的炎症有关。果糖诱导的肝脏脂肪生成还涉及其他菌群代谢物,如乙酸。膳食果糖通过增加小鼠内毒素刺激后的循环IL-1b发挥炎症作用,可能是由果糖依赖性诱导mTORC1介导的。尽管一些临床前研究表明膳食果糖会改变肠道菌群,在10名健康志愿者中进行的小规模人类试点研究却无法建立此关联。
另一个重要的促炎、破坏肠道屏障和影响肠道菌群的饮食成分是酒精。酒精已被确定为扰乱肠道菌群和促进肠-肝轴炎症反应的最关键宿主变量之一。后文会详细展开讨论。
氧化三甲胺 (TMAO)是饮食促进肠-肝轴的另一个例子。肠道细菌将膳食中的胆碱或肉碱(肉类和奶酪中含有)转化为TMA, TMA在肝脏中通过黄素单加氧酶(FMO3)进一步代谢为TMAO。研究表明,这一关键的肠-肝轴生物标志物与系统性疾病,特别是心血管疾病有关联。在活检鉴定的NAFLD中,肝病的严重程度与较高的TMAO水平相关,TMAO水平与T2D中NASH的严重程度相关。尽管许多研究已经揭示了TMAO与疾病预后的密切联系,但其潜在机制尚不清楚。临床前研究表明,TMAO影响胰岛素和葡萄糖耐量,并促进脂肪组织炎症。在此概念下,促炎饮食在肝脏中代谢,相关代谢物控制肝脏内外的免疫,被称为免疫代谢。
抗炎的饮食
健康的生活方式和营养有利于产生抗炎代谢物,然而对于这样的饮食应该是怎样的,仍存在着相当大的争论(部分原因是对营养的异质反应)。尽管脂肪、单糖和大多数蛋白质在小肠中被消化,但复杂的碳水化合物(即纤维)在结肠中被共生菌发酵成SCFAs,以促进机体健康。这些SCFAs与宿主的特定受体结合,例如G蛋白偶联受体(GPCRs),诸如 GPCR41和GPCR43,从而在能量代谢和免疫耐受中发挥有益功能。SCFAs调控调节型T细胞的占比和生物学功能,从而有助于肠道内稳态和预防结肠炎。SCFAs改善脂肪变性和肝脏炎症,并进一步延缓乙肝病毒HBx转基因小鼠慢性肝病向HCC的进展。因此,膳食纤维不仅可以影响肠道菌群落和功能,还可以有效改善实验性代谢性疾病。
目前,已发现的有益的菌群代谢物数量正在增加,芳香烃受体(AhR)配体就是一个突出的例子。许多天然的AhR配体存在,从膳食成分到菌群衍生的代谢物,如色氨酸,与特定受体结合转移至细胞核,并与AhR核转位器结合。这种相互作用影响许多可能抑制炎症活性的基因的表达。还发现,与对照组小鼠相比,AhR肠上皮缺失小鼠在乙醇暴露后表现出更严重的肝损伤,提示了沿肠-肝轴的AhR信号通路的作用。AhR配体在代谢综合征和肝脏脂肪变性的临床前和临床环境中也显示出多效性,可能是通过改善肠道屏障完整性发挥作用。
肠-肝轴中的微生物信号
1. 病原体相关的分子模式
受损的肠道屏障是潜在有害菌及其效应分子(被称为病原体相关分子模式(PAMPs))转移到肝脏的重要前提。内毒素是一种典型的PAMP,是共生菌革兰氏阴性菌细胞壁的组成部分。内毒素反映了肠-肝交流中的一个关键的肠源媒介,能有效地诱导促炎细胞因子的释放,特别是结合TOLL样受体4(TLR4)后巨噬细胞释放的促炎细胞因子。在早期的研究中,观察到ALD和NAFLD中内毒素浓度的增加。在诸如T2D、肥胖或NAFLD等代谢性疾病中,内毒素血症已成为全身炎症的重要因素。内毒素血症与肝病严重程度相关;且与单纯脂肪变性相比,NASH患者的循环内毒素水平更高。
除了内毒素,许多其他PAMP,如脂磷壁酸、肽聚糖、脂肽类或来自病毒和细菌的DNA,都可能进入门静脉,到达肝脏甚至胆汁。健康的肝脏有对抗和清除内毒素的机制。最近,Han等报告肠道衍生的高密度脂蛋白HDL3使内毒素失活,证实了胆固醇的一种新作用。肠源性HDL3可以中和门静脉中的内毒素,防止肝脏巨噬细胞的激活和肝脏炎症,这一点也在手术、饮食或酒精性肠道损伤的动物模型中得到证实,其中肠道HDL3的耗竭会加重肝脏损伤。该研究表明,肠源性因子——HDL3,可以保护肝脏免受炎症的损害,临床慢性肝病可能与此有关。
胆汁淤积实验模型小鼠表现出从窦性血液中清除时间的延迟,管腔内毒素的分泌量减少,这表明BA代谢在调控肝细胞PAMP清除方面的作用。慢性内毒素暴露对慢性肝病内毒素耐受性和/或训练免疫的作用目前尚不清楚。也有报道,防御素等抗菌肽在正常肝脏中显著表达,且在胆汁淤积条件下被BAs和胆红素进一步诱导。
2. 影响肠-肝轴的菌群代谢物
只有少数临床研究阐述了代谢性肝病中的门静脉血、菌群和衍生代谢物。Koh等报道T2D患者门静脉血液中菌群代谢物丙酸咪唑浓度增加,影响胰岛素信号传导,其中丙酸咪唑通过激活p38 MAPK和p62的磷酸化导致mTOR活性增强,从而损害胰岛素信号。McDonald等研究了共代谢物D-乳酸,它能够清除肝细胞中的病原体,在抗生素处理后逆转。因此,菌群代谢物可以调控肝脏免疫反应。
在实验性NAFLD中,共代谢物苯乙酸引起肝脏脂肪变性,将肥胖妇女的粪便转移到小鼠中导致肝脏脂肪变性加重。同样,菌群代谢物N,N,N-三甲基-5-氨基戊酸(TMAVA)可能在肠-肝轴上发挥作用,因为NAFLD患者血清中,TMAVA水平升高。总的来说,还需要更多的机制研究来了解门静脉代谢物如何影响小鼠和人类的代谢性肝病。
肠粘膜激素:FGF15/19和GLP1
除了饮食和微生物信号外,宿主衍生的介质也参与肠-肝轴。例如,“肠因子”,如小鼠中的成纤维细胞生长因子15 (FGF15)(对应人类中的FGF19),主要由回肠的肠道上皮细胞产生,而不是由肝脏产生。FGF15/19在回肠中的表达主要受胆汁酸激活的法尼醇X受体(FXR)的转录控制,构成肝-肠轴。FGF15调控BA合成和肝脏糖脂代谢,因此,FGF15参与了沿肠-肝轴的双向交流。而且,FGF15/19也是一种关键的餐后葡萄糖和能量代谢调节剂,通过抑制CREB-PGC-1a(环磷酸腺苷- (cAMP)反应元件结合过氧化物酶体增殖物激活受体γ共激活因子1- α)信号通路影响糖异生,而FGF15转基因小鼠表现出更高的代谢率和更低的葡萄糖和胰岛素浓度。尽管已报道其对脂肪组织和大脑的影响,肝脏仍是FGF15/19的关键靶器官。FGF15/19临床应用的主要难点是其在肝脏上的有丝分裂和潜在的致癌活性。这可以通过构建工程突变体FGF19-M70来克服,该变体影响BA代谢,但缺乏有丝分裂活性。FGF19-M70 (NGM282)的临床试验目前已在原发性胆道胆管炎(PBC)、原发性硬化性胆管炎(PSC)和NAFLD等肝脏疾病中进行。特别是,在一项II期临床试验中,这种化合物能够减少NAFLD的肝脏脂肪和纤维化,提示回肠激素可能改善肝脏疾病。
胰高血糖素样肽1 (GLP1)是回肠和结肠中的特化上皮细胞分泌的一种肠道激素,控制肝脏中的葡萄糖代谢和胃排空和饱腹感。GLP1受体激动剂用于T2D患者的高血糖治疗,并可改善心血管预后。目前,GLP1激动剂被探索用于治疗人类肝病(在有效的阶段性临床试验中),因此,利用肠-肝轴进行治疗干预可能成为未来NAFLD患者的主要治疗方法。GLP1激动剂的有益作用可能是通过减轻体重和控制糖代谢介导的。
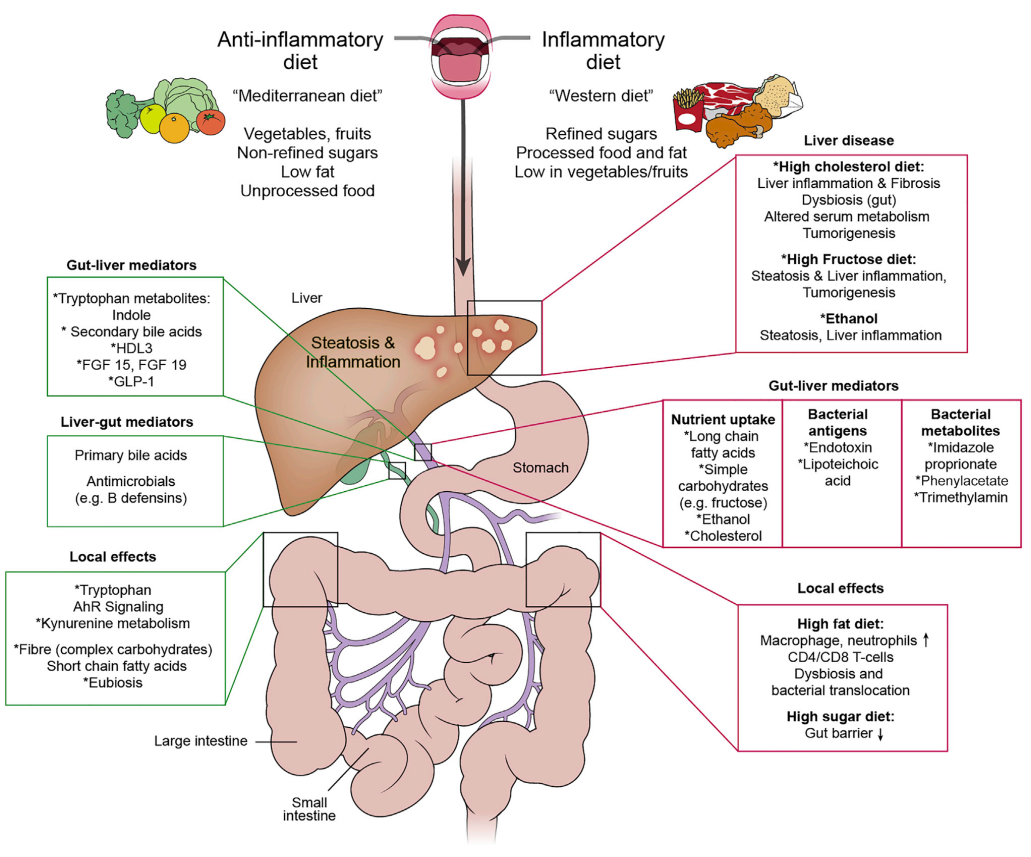
图2. 在健康和疾病中肠-肝轴双向交互的介质
参考文献
Gut-liver axis: Pathophysiological concepts and clinical implications. Cell Metabolism. 2022.
07-01 英斯特朗
连载 | 药物一致性评价与粒度分析(三)07-01 欧美克仪器
【仪器百科】LS-909丨干湿二合一激光粒度分析仪07-01 欧美克仪器
标准物质解决方案 | PFASs(全氟及多氟化合物)06-29
第九期阿尔塔有约 | 环境专题【新污染物:PFAS】技术研讨会精彩回顾及提问解答06-29
“绿色技术范式”,分析化学未来发展方向——访中国分析测试协会副理事长、辽宁省分析科学研究院原院长刘成雁教授06-29 转载仪器信息网
华西医院-标准型数显脑立体定位仪、双通道体温维持仪、体式显微镜安装完成06-29 迈越生物
科鉴检测助力2家仪器企业获得首批产品可靠性认证证书06-28 科鉴检测
德国耶拿:锂电池生命周期分析解决方案06-28 德国耶拿
AI已来!生命科学本科教学如何紧跟技术浪潮06-28 Opentrons
盛瀚售后,五星级服务的秘诀是什么?06-28 SHINE
专为汽车制造商打造的柔性解决方案——实现制程控制06-28
西北工业大学-脑立体定位仪安装完成06-28 迈越生物
会议邀请 | 第九届海上检验医师论坛06-28
卓立要闻 | 创新发展ing…6月卓立“大事小情”速览06-28 光电行业都会关注
打造信任合作伙伴!2024年度卓立汉光客户满意度调查开启06-28 光电行业都会关注
如何挑选适用于三阶光学非线性的测量系统?Z扫描测量系统来助力!06-28 光电行业都会关注
招聘启事—中国科学院沈阳自动化研究所微纳光学测量表征技术课题组06-28 光电行业都会关注
谱育科技作为主要完成方 荣获2023年度国家科学技术进步一等奖和二等奖06-28 点击关注→
仪器原理丨顶空仪与吹扫捕集仪科普小知识06-28 天美色谱