项目文章Plant Physiol (IF 8.34)| 喻德跃教授团队采用磷酸化组揭秘大豆开花和抗虫之间的协调关系
2022-06-27 11:18:53, APT-MKT 上海中科新生命生物科技有限公司

光周期敏感的植物,如大豆,在整个生长期都面临食叶性昆虫的危害,而开花期经常与害虫高发期一致。目前,尚不清楚植物如何协调其开花和抗虫性之间的关系。
2022年5月31日,国际顶尖植物期刊Plant Physiol(IF 8.34)在线发表了南京农业大学喻德跃教授团队李霄等老师的创新研究成果“CALCIUM-DEPENDENT PROTEIN KINASE38 regulates flowering time and common cutworm resistance in soybean”。该研究揭示了大豆GmCDPK38在调控开花时间和抗虫性中的双重功能及其在大豆驯化中的进化规律,为植物开花与抗虫性的研究方向提供了新思路。其中中科新生命提供了磷酸化蛋白质组检测服务。

研究材料
大豆Jack(WT植株)、gmcdpk38突变体和携带GmCDPK38 Hap2或Hap3单倍型的大豆材料
技术路线
步骤1:光周期调控GmCDPK38的表达;
步骤2:GmCDPK38的遗传变异与大豆开花时间和对斜纹夜蛾(CCW)的抗性相关;
步骤3:CRISPR/Cas9介导的GmCDPK38 Hap3靶向突变导致大豆开花延迟,并增强对CCW的抗性;
步骤4:利用磷酸化蛋白质组筛选GmCDPK38的下游底物;
步骤5:RNA-seq分析揭示GmCDPK38影响的基因表达;
步骤6:gmcdpk38突变体中多种代谢产物的积累增加;
步骤7:GmCDPK38翻译后调节S-腺苷甲硫氨酸合酶;
步骤8:GmCDPK38是一个驯化基因,抗性的Hap2可能是一个人工选择靶点。
研究结果
1. 光周期调控GmCDPK38的表达
GmCDPK38是钙依赖性蛋白激酶第IV亚家族成员。通过光周期实验发现GmCDPK38在长日照(LD)条件下的表达水平大于短日照(SD)。随后通过分析GmCDPK38的日表达模式,发现在LD条件下,GmCDPK38的表达量在白天减少,夜间增加。在SD条件下观察到该基因存在相同的表达模式,但其转录的幅度和丰度低于LD。这些结果表明,GmCDPK38的表达受光周期调节,在LD条件下表达量更高(图1)。
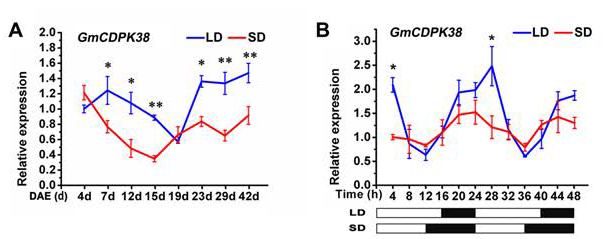
图1 GmCDPK38表达分析
2. GmCDPK38的遗传变异与大豆开花时间和对斜纹夜蛾(CCW)的抗性相关
分析了272株栽培大豆中的GmCDPK38序列,在至少7种环境中,18个多态性位点与开花时间的变化显著相关。其中,含单倍型2(Hap2)的大豆的开花时间明显晚于含Hap3。此外,具有晚花Hap2的栽培大豆对CCW更具抗性。在LD和SD条件下,Hap2中的GmCDPK38表达显著低于Hap3,其启动子活性也低于Hap3。综上,GmCDPK38可能在花期和抗虫性方面发挥作用(图2)。
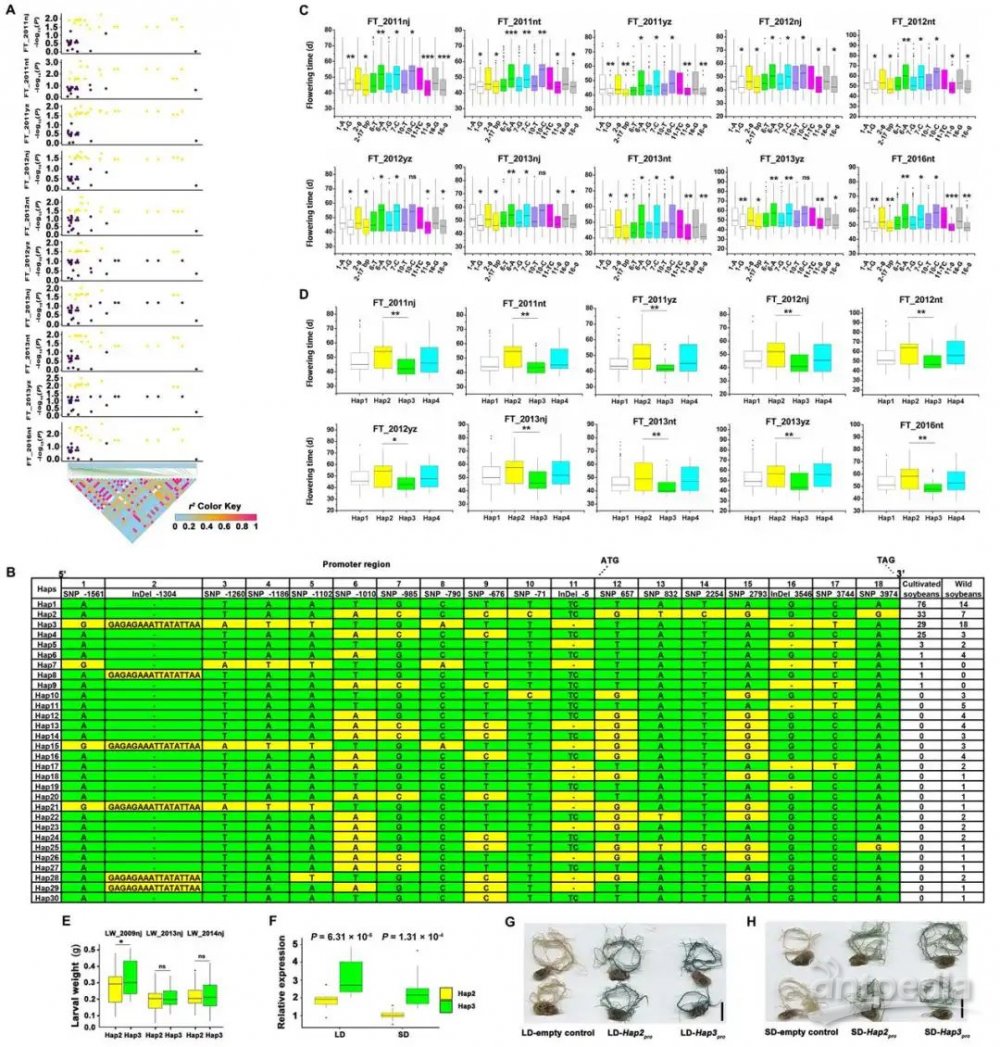
图2 大豆GmCDPK38单倍型分析
3. CRISPR/Cas9介导的GmCDPK38 Hap3靶向突变导致大豆开花延迟,并增强对CCW的抗性
大豆品种Jack中的GmCDPK38序列属于Hap3。比较野生型(WT)Jack植株和gmcdpk38突变体的开花时间,发现突变体的开花时间晚于WT植株。此外,在LD条件下评估gmcdpk38突变体和WT植株的抗虫性,实验表明GmCDPK38的突变提高了大豆对CCW的抗性(图3)。

图3 T3 gmcdpk38突变体的表型鉴定
4. 利用磷酸化蛋白质组筛选GmCDPK38的下游底物
通过定量磷酸化蛋白质组学分析检测WT植株和gmcdpk38突变体中的磷酸蛋白。数据显示了一系列可能的GmCDPK38激酶下游底物,推断GmCDPK38可能通过影响这些蛋白质的磷酸化来介导大豆开花和抗虫性(图4)。

图4 磷酸蛋白质组数据总结
5. RNA-seq分析揭示GmCDPK38影响的基因表达
通过RNA-seq鉴定了大量与抗性和开花相关的表达基因。GmCDPK38突变诱导大多数开花抑制基因的表达,而gmcdpk38突变体中大多数开花促进基因表达下调。此后通过RT-qPCR证实这些基因表达与RNA-seq结果一致(图5)。

图5 GmCDPK38诱导T3 gmcdpk38突变体基因表达的变化
6. gmcdpk38突变体中多种代谢产物的积累增加
对WT和gmcdpk38突变体进行靶向代谢组学分析。在LD下42天,与WT相比,突变体中65.28%的代谢产物含量显著增加,其中包含16个不同的化合物类别。大多数差异代谢产物,包括类黄酮、苯丙烷、多酚、酚酰胺、生物碱和萜类,都是已知的植物内源性防御代谢物。SD下29天的代谢物水平下降的比例(48.93%)高于LD下42天。综上,GmCDPK38的突变诱导多种代谢物的积累,尤其是在LD条件下(图6)。

图6 T3 gmcdpk38突变体各类差异代谢产物数量
7. GmCDPK38翻译后调节S-腺苷甲硫氨酸合酶
磷酸化蛋白质组学数据显示S-腺苷甲硫氨酸合酶GmSAMS1仅在WT组中发生磷酸化。通过酵母双杂交(Y2H)和双分子荧光互补(BiFC)实验,发现GmCDPK38与GmSAMS1互作并可能导致GmSAMS1磷酸化。进一步研究发现与防御相关的S-腺苷甲硫氨酸合酶在gmcdpk38突变体中在翻译后上调表达,表明GmCDPK38突变增强了大豆对CCW的抗性(图7)。

图7 T3 gmcdpk38突变体S-腺苷甲硫氨酸合酶翻译后上调
8. GmCDPK38是一个驯化基因,抗性的Hap2可能是一个人工选择靶点
Hap2在栽培大豆中出现的频率高于野生大豆,多出现在低纬度地区,而Hap3在野生大豆中出现的频率更高,主要分布在高纬度地区。Hap2在主栽品种的高频出现表明该等位基因可能是人工驯化的结果。通过计算驯化指标,发现GmCDPK38在大豆驯化中受到选择(图8)。

图8 GmCDPK38在大豆驯化中受到选择
小编小结
本研究利用基因编辑、磷酸化修饰组学、代谢组学等分析证实钙依赖性蛋白激酶GmCDPK38在协调大豆开花时间调节和抗虫性方面起着双重作用。GmCDPK38在野生大豆中具有丰富的遗传多样性,可能是在大豆驯化过程中选择的。

中科优品推荐
【中科新生命】不仅提供磷酸化蛋白质组,也提供磷酸化蛋白组+蛋白组学的多组学检测及联合分析服务,助力深度挖掘激酶与底物磷酸化的调控关系,多维度剖析调控机制。
中科新生命为您提供最完整、最可靠的组学技术服务!


<< 滑动查看下一张图片 >>


我知道你
哦

10-24 莲莲看
VELP唯意朴仪器|为环境保护提供关键分析仪器和实验室设备10-24
惠州学院八台热分析仪安装调试完毕,助力科研教学再上新台阶10-24
伊萌检测技术服务有限公司选购我司HS-DSC-101差示扫描量热仪10-24
鲁东大学选购我司差示扫描量热仪10-24
差示扫描量热仪工作原理是什么?10-24
和晟仪器亮相2023碳中和国际学术会议10-24
厦门市宜帆达新材料有限公司选购我司HS-DSC-101差示扫描量热仪10-24
今日霜降丨气肃而凝,露结为霜10-24
天美讲堂丨光学显微镜中的照明技术10-23 天美
美谱达仪器:助力技能大赛,成就技能人才10-23 Mapada
Quantum Design中国用户科研成果快报(2023年第8期)10-23 Dr. Sun
Quantum Design中国用户科研成果快报(2023年第7期)10-23 Dr. Sun
直播预告丨物性测量“沙拉Jiang”系列讲座之镍基超导、vdW铁磁材料、热电材料研究10-23 Dr. Sun
相约庐州|2023全国流行病学学术会议10-23
重阳 | 群山之巅 登峰造极,每一步都是新高度10-23 SHINE
聚焦天隆|天隆智造再次上榜“优秀国产医疗设备目录”10-23
重阳节丨登高望远处,美好踏秋时10-23
会议邀请|精细化工全生命周期安全风险评估技术交流会(济南站)10-23
GLMY创想仪器丨参加2023潍坊铸造展10-23 国产精密仪器厂家