Nature Metabolism | 为乳酸正名:能量代谢中的丑小鸭
2020-08-14 02:06:02, Volibear 麦特绘谱生物科技(上海)有限公司

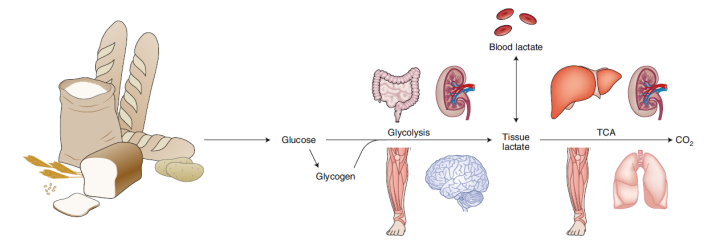
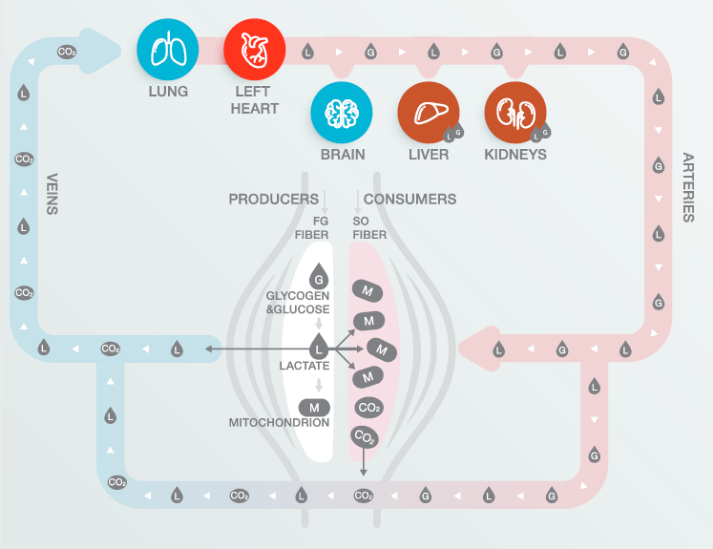

参考文献

解锁更多精彩「代谢组学」相关资讯!
微信公众号:麦特绘谱
Tel:400-867-2686
Web: www.metaboprofile.com
识别二维码,关注我们
Volibear 的近期文章
其他厂商的近期文章
HC来啦!衡昇质谱期待与你同行
01-24 衡昇质谱
华枝春满四十载 携手前行铸辉煌01-23
2024年国家自然科学基金【岩土方向】|纽迈低场核磁应用解决方案01-23 纽迈分析
GE医疗驰援海东地震灾区医疗设备已在医院就位01-22
重磅!全新Ergo KPFM让表面电势的测量简单又准确01-22 牛津仪器
屹尧科技助学金,温暖莘莘学子心01-22 屹尧科技
助力双碳,“氢”心打造-燃料电池汽车用氢质量分析方案(Ⅰ)01-22 飞飞
前沿应用|低场核磁共振技术用于固体药物多晶型的定性与定量研究(二)01-22 纽迈分析
岛津分析客户服务中心升级启航一AI引领向着数字化和自动化演进01-22 仪器信息网
1月23日直播 | 微观与宏观的美丽邂逅——先进金属材料前沿表征技术研讨会01-22 分析中心
哈希京东旗舰店店铺会员日01-22
偏振分光镜产品介绍01-22 韵翔光电
大寒却暖雪晴天01-21
2023元析仪器年会盛典 | 突破·创新,析望无限。01-20 元析仪器
寒气之逆极,故为大寒01-20
擦亮品牌 凝聚力量 | 皖仪科技获“行业先锋品牌”称号01-19 WAYEAL皖仪
【新功能】非靶向代谢组学中的同位素示踪01-19 核磁同位素学社
有奖互动 #谁最爱用过柱机?01-19 市场部
三度蝉联!丹纳赫荣膺“中国杰出雇主”认证01-19
惊!都2024了 还有人不知道儿茶酚胺怎么分析嘛01-19 王静