Cancer Cell | 肿瘤学一区顶刊:空间多组学技术破译胶质母细胞瘤中的肿瘤-宿主相互依赖性
2022-07-18 07:40:26, 多层组学定制服务 上海欧易生物医学科技有限公司


(点击图片报名千万医学支持计划)
前言
单细胞技术为健康人脑和恶性肿瘤中单个细胞的转录调控和动态进化提供了见解。神经胶质瘤(GBM)的单细胞RNA 测序(scRNA-seq) 研究表明,肿瘤内异质性和跨细胞状态的动态可塑性是恶性脑肿瘤的标志。尽管脑肿瘤呈现出与健康大脑相似的转录适应和进化,但它们长期以来一直作为一个独立的实体进行研究,忽略了局部微环境的作用在肿瘤发生中。由于空间组织信息的丢失,单细胞分析仅提供细胞相互作用的间接推断。空间转录组学是一项新技术,它使作者能够原位表征细胞相互作用和组织,从而破译恶性脑肿瘤的生态系统。
2022年6月,来自德国弗莱堡大学医学中心的Dieter Henrik Heiland团队在Cancer Cell期刊(IF:38.585)发表的题为“Spatially resolved multi-omics deciphers bidirectional tumor-host interdependence in glioblastoma”的研究成果,采用空间转录组学、空间代谢组学和蛋白质组学(成像质谱流式,IMS)研究方法,发现了胶质母细胞瘤基本特征,并确立了以免疫和代谢应激因素为标志的微环境,阐述了局部区域肿瘤-宿主的相互依赖性。这些研究进一步强调了在空间分辨的背景下由于微环境引起的神经胶质瘤的各种转录适应的必要性,为进一步探索早期胶质母细胞瘤复发和治疗抗性提供了重要信息。

研究背景
鉴于 scRNA-seq 和空间转录组学的互补性,这两种技术的整合至关重要。除了空间分辨的转录组外,还需要补充分子分析来全面了解微环境对肿瘤细胞的影响。细胞通讯和新陈代谢是对脑癌的动态适应、促进生长、浸润和治疗抵抗具有决定性影响的两个关键因素。
本研究进一步强调了在空间分辨的背景下由于微环境引起的神经胶质瘤的各种转录适应的必要性。旨在提供胶质母细胞瘤与其局部微环境的空间分辨转录程序和细胞相互作用的图谱。
研究思路
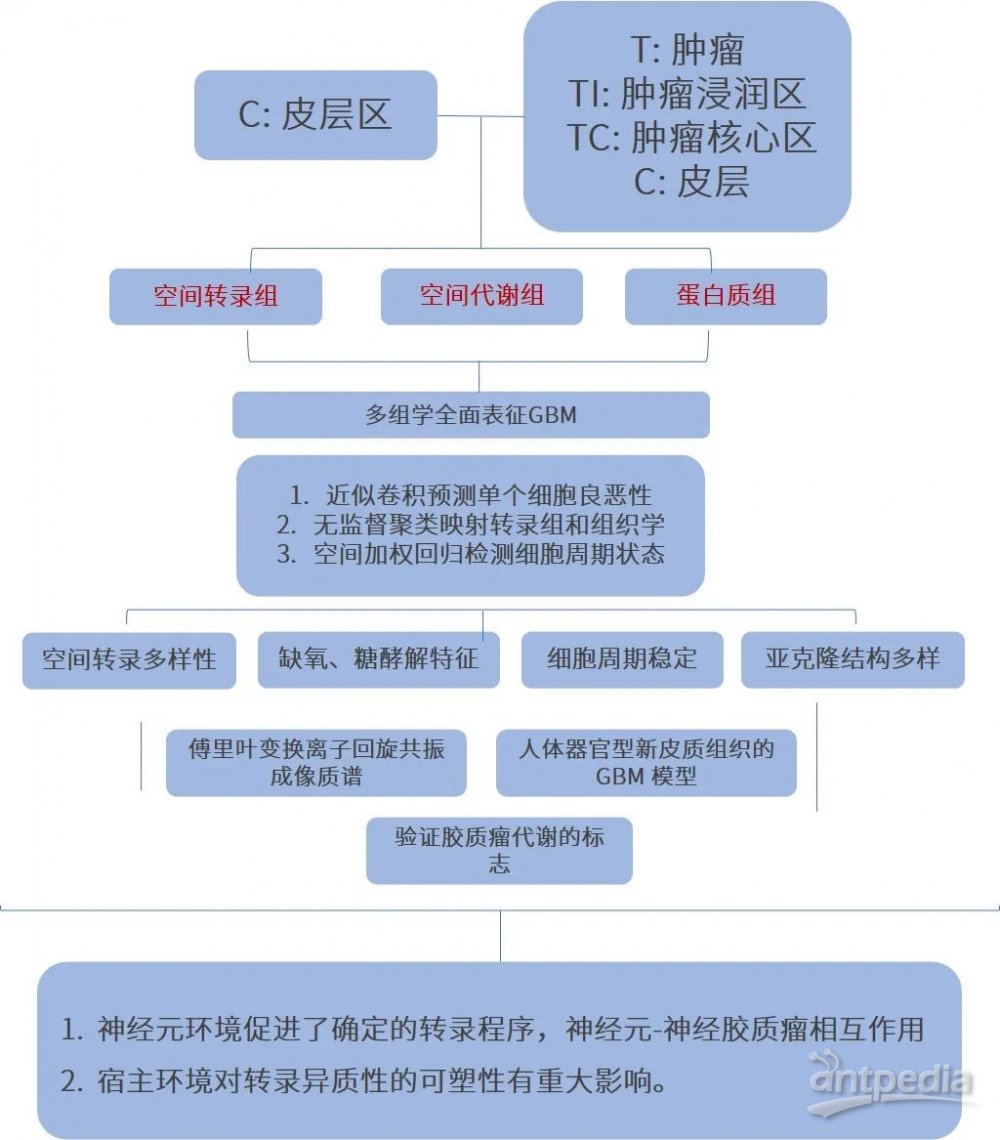
研究结果
1.胶质母细胞瘤空间分辨转录组学图谱
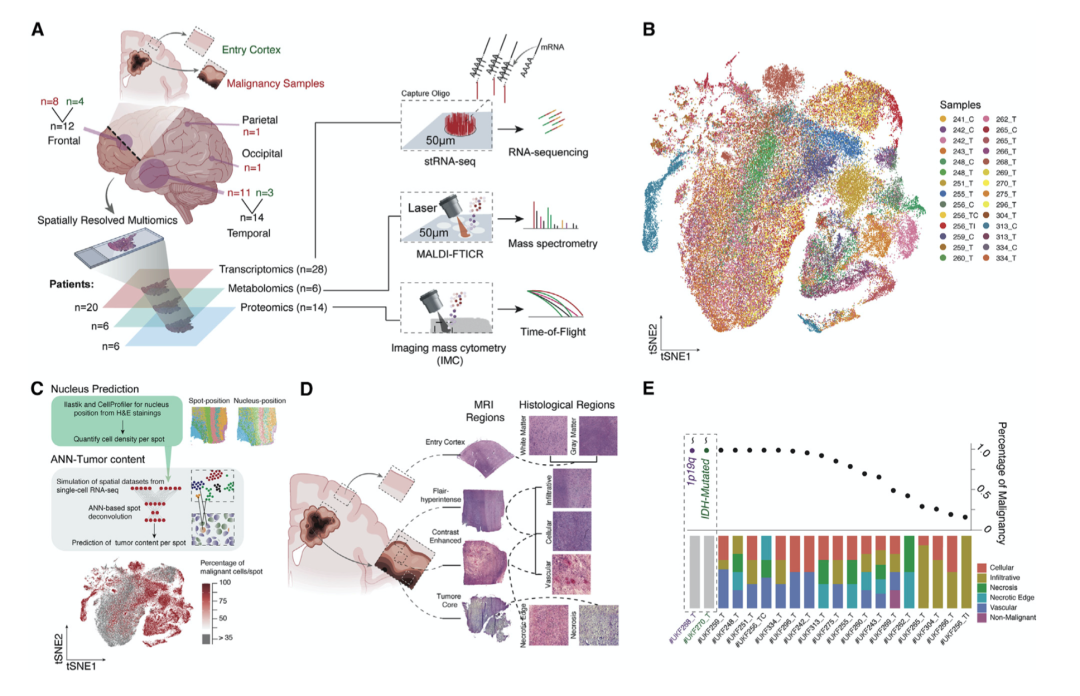
为了表征胶质母细胞瘤(GBM) 的空间结构,作者选取了 28 个样本(患者 n = 20)的空间分辨转录组学(stRNA-seq)图谱,产生了 88,793 个跨不同年龄组和解剖学的个体转录组。作者还补充了来自组织的空间分辨代谢组学(来自患者 n = 6)和蛋白质组学(来自患者 n = 6的 n = 14 个感兴趣区域 [ROIs] )用于 stRNA-seq 辅助整合,以全面整合多个分子层(图 1A)。
基于相互最近邻 (MNN) 的水平整合和共享最近邻 (SNN) 聚类显示,非恶性标本在患者之间表现出相似性。恶性转录组以独特的基因表达谱为标志,这进一步证实了跨簇样本多样性的指数(图 1B)。恶性样本的指数降低,表明这些簇由来自个体患者的斑点组成(图 1B和 1C)。这些发现与最近的单细胞研究一致,即大量个体拷贝数改变和突变谱有助于患者间的异质性,从而导致个体转录谱改变。
在所有样本的 88,793 个点中,63,121 个点来自恶性标本,其中 46,459 个点包含至少 95% 的肿瘤细胞(图1C)。为了研究浸润区域,作者根据 Ivy GAP 组织学分类系统预测了逐点组织学表型(图 1D)。与组织病理学特征相比,具有低肿瘤细胞频率的样本主要包含浸润区域,在这些区域中,健康皮层内的肿瘤细胞难以描绘(图 1E)。
2.破译空间分辨的转录异质性
通过与现有的转录和组织学分类数据联系来重现空间转录模式,进一步确定GBM 转录亚群的空间定位。运用空间转录组学,作者发现了五个一致表达的转录程序,后来被称为“空间不同的转录程序”,其特点是每个程序在 90% 的调查肿瘤中都存在(图2A)。
作者随后整合了患者空间加权相关矩阵(逐个聚类),然后是层次聚类,确认了五个重复的空间不同转录程序的空间分离。图2A)。这种方法使作者能够了解在整个队列中稳健表达的患者特异性簇中转录程序的生物学意义。转录程序与神经胶质谱系显示出实质性的一致性,并根据它们的神经(称为“神经发育”)或少突胶质细胞起源(称为“空间 OPC”)命名, NPC 和 OPC 样肿瘤细胞亚组(图 2 C)。
为了进一步将作者的发现与现有的分类系统相结合,作者对已建立的体积进行了空间加权回归,以及作者在空间上不同的转录程序分类来映射空间关系(图 2C)。与 Richards 等人的数据相比,上述所有程序都显示出与神经发育表型的强烈重叠(图2C)。反应性缺氧计划与 MES 样亚型表现出最强的重叠,特别是缺氧依赖性“MES2”状态(图2C)。
拷贝数改变 (CNA),包括癌基因的局灶性扩增或肿瘤抑制因子的丢失,是恶性细胞的标志,据报道会导致治疗耐药和肿瘤复发。在已知的细胞状态分类中独特的 CNA 与特定状态相关联,在单个肿瘤细胞中不均匀分布。作者利用推断的 CNA 研究了基因组改变和空间上不同的转录程序之间的关系,并揭示了重要的阈值以增加作者推断方法的稳健性。在与反应性缺氧相关的点中,作者发现了作为独立亚克隆事件发生的 CNA 的显著积累(图 2 D)。

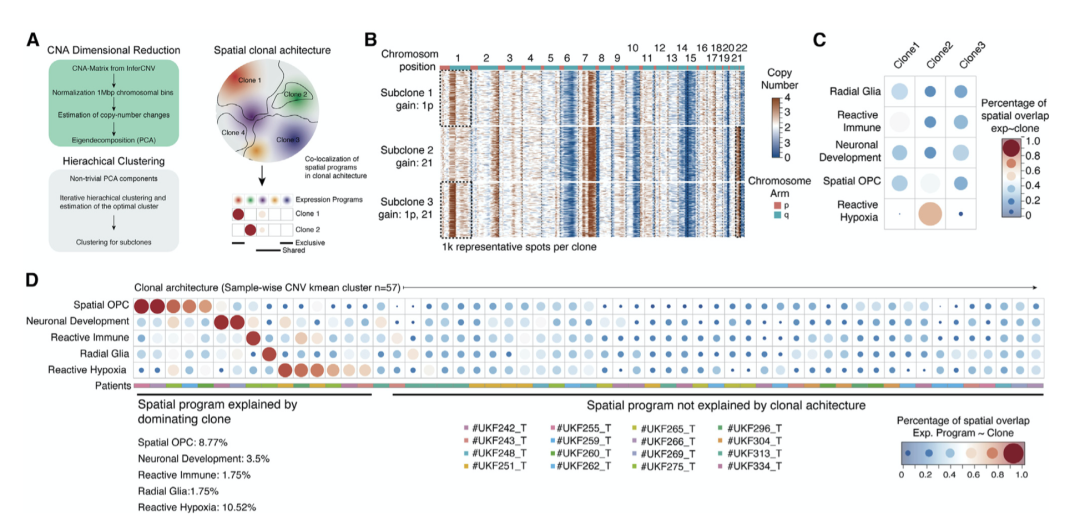
3. 空间上不同的转录程序和亚克隆结构
作者进一步研究空间上不同的转录多样性是否可以直接反映肿瘤内的遗传亚克隆。作者通过 CNA 的患者特异性层次聚类重建了克隆体系结构。鉴定了 57 个遗传亚克隆,每个样本跨越 2 到 6 个亚克隆(图 3A)。然后,作者确定了每个亚克隆中个体、空间上不同的转录程序的分布(图 3B 和 3C)。分析表明,在 26.32% 的所有亚克隆中,单一转录程序占主导地位。这些发现帮助作者得出结论,尽管亚克隆偶尔偏向于空间 OPC (8.77%) 或反应性缺氧程序 (10.52%),但克隆结构对空间不同转录程序的发生影响有限(图3D)。
4. GBM中与反应性缺氧程序相关的代谢改变
为了进一步探索空间上不同的反应性缺氧模式,作者使用空间代谢组学技术从连续用于 stRNA-seq 的组织切片(患者,n = 6) (图 4A )。实施了一个类似于 stRNA-seq代谢谱的工作流程,降维揭示了前两个主要成分中的主要差异,从而确定了三个显著的代谢亚组后校正集群稳定性(图 4A、4B)。功能代谢分析显示在第一代谢模块(M-G1)磷酸戊糖途径显著富集。第二个代谢亚组(M-G2)的特征是磷酸腺苷酸代谢富集。最终的代谢亚组(M-G3)在糖酵解和氨基糖代谢方面高度富集(图4C )。
针对空间上不同的转录程序反应性缺氧,在作者的综合分析(WNN 聚类)中作为一个独特的簇出现(图4B)。组织学上,分配给反应性缺氧程序的大多数斑点代表坏死边缘(图 4 C)。作者假设低氧代谢导致基因组不稳定性在空间隔离的生态位中积累。这些“反应岛”代表了从头的潜在来源基因组改变有助于肿瘤细胞产生治疗抗性。
进一步探索富含反应性缺氧特征的区域。区域被定义为缺氧核心,并富集糖酵解途径(图 4 D)。还显示出多条染色体(8p、9p、13q、19q 和 21q)的独特损失和增加,这种变异最有可能由缺氧核心区域内的单个亚克隆解释,这证明了缺氧相关代谢是一种潜在的假设基因组不稳定性的驱动因素(图 4 E)。

5. 反应性缺氧区域积累染色体改变
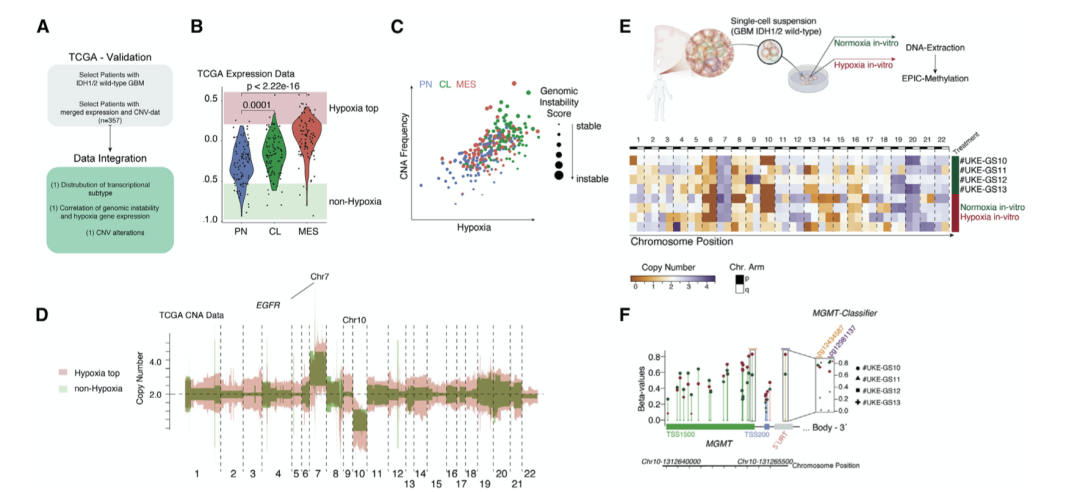
作者分析了来自 TCGA 数据库的样本(患者 GBM IDH1/2野生型 n = 357),根据患者的缺氧基因表达评分对患者进行分类(图 5A和 5B )。缺氧驱动的肿瘤显示染色体改变显著增加,证实了代谢和基因组不稳定性之间的关系(图 5C 和 5D)。结果主要表明,低氧/氧化应激增强了基因组的不稳定性,有助于在亚克隆选择的支持下加强 GBM 的微进化以恢复弹性。为了通过实验验证作者的假设,作者在常氧和缺氧条件下培养原发性患者来源的GBM 细胞系2-6 周(患者 n = 4)(图 5E)。拷贝数变异揭示了 CNA 谱的差异,在慢性缺氧条件下 CNA 事件的显著积累证实了压力诱导的 CNA 改变的假设(图 4 E)。
作者通过分析常氧和缺氧条件下CpG 位点cg12434587 和 cg12981137处的 O-6-甲基鸟嘌呤-DNA 甲基转移酶 (MGMT) 启动子甲基化来探索缺氧代谢对DNA 甲基化的影响。细胞系当暴露于缺氧时,在常氧下具有未甲基化的 MGMT 启动子会产生高甲基化(图 5 F)。这些发现进一步加强了关于代谢驱动的 CNA 的假设,其中大多数从头重排必须被视为随机事件,有助于肿瘤的亚克隆进化。
6. 低氧代谢调节GBM中的“去或成长”潜力
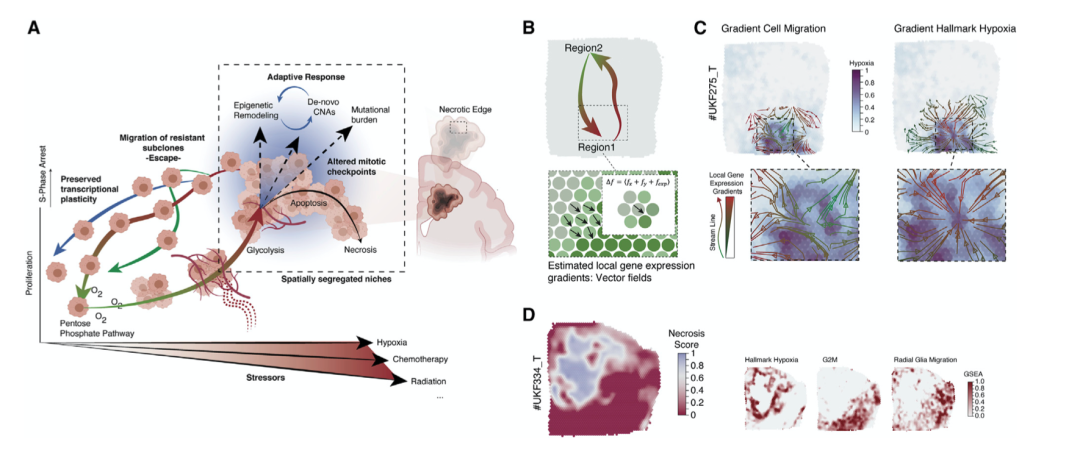
在对空间分辨转录程序的初步表征中,反应性缺氧程序在非循环细胞中显著富集(图2A)。表明缺氧应激会触发细胞周期停滞,特别是 S 期停滞(图 6A)。导致假设缺氧驱动的 S 期停滞有助于基因组不稳定性的积累(图6A )。
GBM 细胞的基线转录状态可以分配给“发育”(AC-、OPC-或 NPC 样)程序。在空间上,代谢谱显示磷酸戊糖途径 (PPP) 的富集。由于增殖和肿瘤生长,会发生越来越多的营养和氧气缺乏,迫使采用代谢程序进行糖酵解。这种现象是 PPP 和糖酵解之间的相互转换,与肿瘤细胞的生长或生长潜力有关,表明 PPP 支配着由生长的肿瘤细胞组成的区域。
当缺氧发生时,这种低氧诱导的糖酵解转换随后的诱导,这有助于细胞逃逸/迁移到含氧量正常的区域。由于在缺氧条件下发生的细胞周期程序的下调,会发生 S 期停滞,导致随后从头CNA 的积累。作者假设这些受缺氧影响的细胞中有很大一部分随后进入凋亡状态,导致在 GBM 中看到的特征性坏死。只有一小部分细胞通过迁移相关转录程序的上调成功逃脱。除缺氧外,据报道,其他压力源(如放射或化疗)会导致 GBM 出现这种与压力相关的扰动(图 6A)。
根据这个模型,可以看到低氧代谢和细胞迁移之间的反比关系。为了探索代谢改变区域中迁移基因表达特征的富集,作者确定了特定基因表达特征的低富集和高富集之间的定向梯度的空间方向。每个点的方向向量是基于其局部邻域中所研究的基因表达特征的分级富集。这些向量场计算使作者能够近似空间基因表达轨迹,从而能够识别空间上相反的转录途径(图 6B)。基于这些矢量场计算,作者报告缺氧响应和迁移特征显示反向空间轨迹(图6C 和 6D)。总之,作者的研究结果为代谢变化和氧化应激是基因组多样性的潜在互惠驱动因素提供了证据,从而导致 GBM 中的克隆进化。
7. 探索反应性免疫区域的肿瘤-宿主相互依赖性
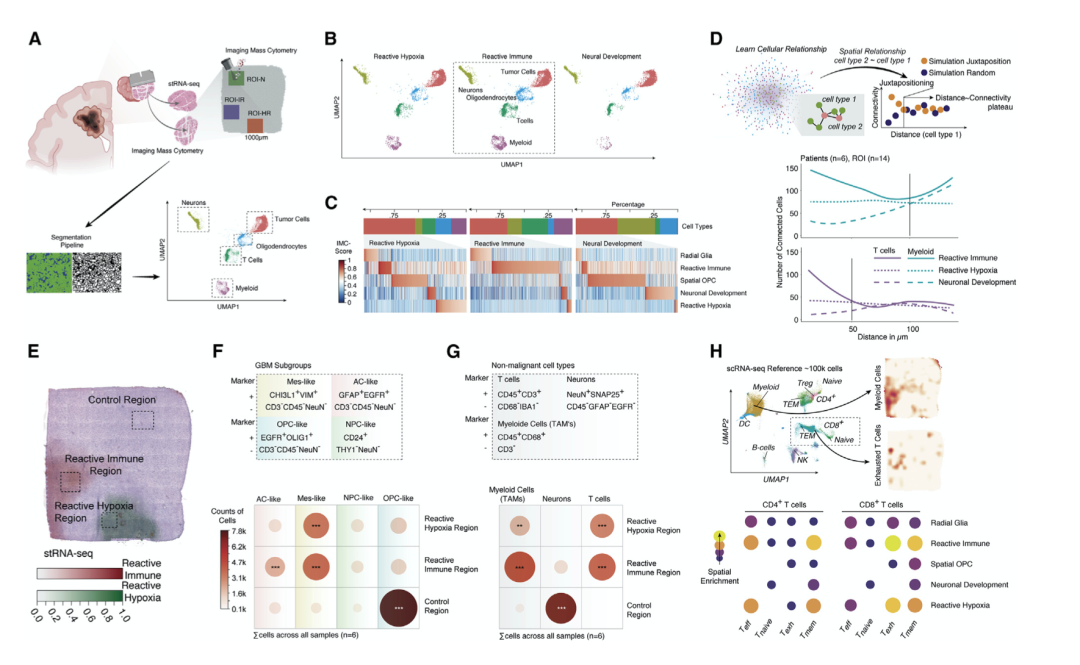
为了提供肿瘤-免疫相互依赖的整体观点,作者用基于单细胞分析成像质量流式细胞术来补充说明。作者选择了反应性免疫和反应性缺氧计划区域内的 ROI,并从分类为神经元分化的区域中控制 ROI 。IMC 数据是从连续用于 stRNA-seq 的组织切片中获取的(n = 6 名患者,14 个不同的 1,000 μm 2 ROI),从而得到分割后 82,179 个细胞的综合蛋白质组图(图 7A;表 S4)。
作者的分析显示骨髓细胞和淋巴样细胞在归类为反应性免疫的区域中(图 7 B、7C)。为了研究免疫富集区域的肿瘤细胞分化,作者通过将细胞细分为放射状胶质细胞(EGFR + HOPX +)、反应性免疫(EGFR + CHI3L1 + VIM +)、空间 OPC(EGFR + OLIG1 + )、神经发育(EGFR + SNAP25 + CALM2 + ) 和反应性缺氧 (EGFR + VEGFA + )。反应性免疫(EGFR)的独家富集+ CHI3L1 + VIM + ) 细胞仅在转录定义的反应性免疫定位中得到证实(图 7 B)。
为了补充关于免疫系统和 GBM 之间细胞关系的发现,作者根据肿瘤细胞与淋巴或骨髓细胞之间的距离量化了它们之间的细胞连接性 ,证实了肿瘤细胞与转录定义的反应性免疫区域中的免疫区室之间的细胞相互作用增强(图7 D)。接下来,作者探索了不同 ROI 中细胞类型的分布。作者证实了在反应性免疫和缺氧中疑似富集 AC 和 MES 样细胞。OPC 样细胞主要富集于神经发育ROI (图 7 E 和 7F)。此外,反应性免疫和缺氧区域均显示肿瘤相关骨髓细胞 (TAM) 和 T 细胞的显著富集(图 7 G)。
鉴于 T 细胞在两个反应区域都富集,作者研究了 T 细胞上的平均PD-1 蛋白水平 (CD3+ ),与反应性缺氧和神经发育相比,反应性免疫区域内显著增加,表明反应性免疫 ROI 中的局部免疫抑制增强。此外,代表 GBM 免疫景观的 scRNA-seq 数据集证实了肿瘤反应性免疫区域中记忆和耗尽的 T 细胞的富集(图 7H)。
8. 环境条件有助于双向亚型转变
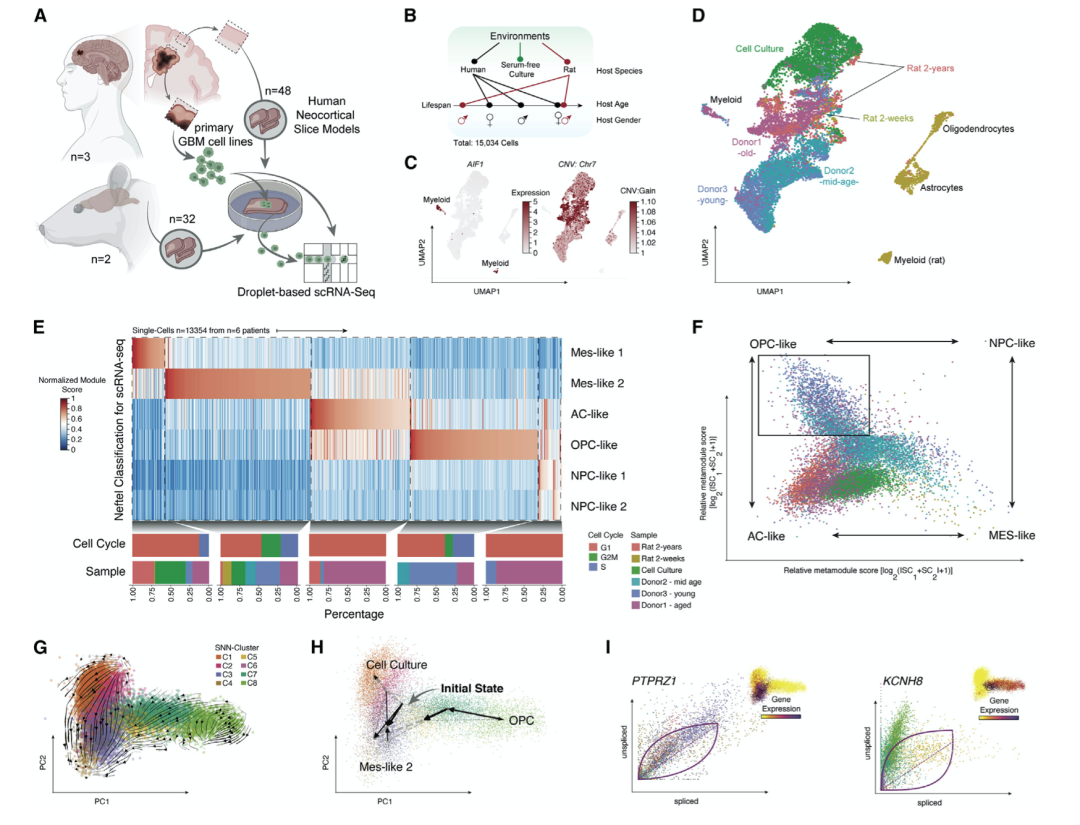
通过之前建立的基于人体器官型新皮质组织的 GBM 模型来模拟了组织浸润,没有代谢和免疫压力。为了评估各种微环境的影响,作者使用了来自不同年龄的多个人类供体(2-85 岁,患者 n = 3;每个患者的切片 n = 48,16)和来自 2 周的两个小鼠供体的皮质通路组织和 2 岁大鼠(n = 32 节)(图 8 A)。将组织培养 4 天,之后相同的 MES 样原发性患者衍生细胞系被接种到所有培养的组织切片中(图 8A)。在不同宿主环境中培养 7 天后,消化组织,并使用 FACS 分离肿瘤细胞,使用 scRNA-seq(15,034 个细胞)进行转录组学分析。使用推断的 CNA(图8C 和 8D),基于肿瘤细胞在 7 号染色体(10,887 个细胞)中的特征性增益进行肿瘤细胞的计算鉴定。
为了探索跨不同宿主环境的动态适应,所有恶性细胞都根据其相应细胞状态进行富集(图 8E 和 8F)。基线状态显示出类似 MES-1/2 的表型,转录多样性程度低且数量众多循环细胞(图 8 E)。主要是在中青年供体中,发育状态在人类供体组织中培养的细胞中显著过多。在啮齿动物神经环境中培养的 GBM 细胞表现出 MES-1/2 和 AC 样转录特征的富集。为了检查动态适应,作者根据前两个主要成分注释了 scRNA速度(图 8 G)。
一般来说,从体外细胞培养(图 8 G 和 8H,顶部)到 MES 和 AC 样,表示由神经环境驱动的方向适应。蛋白质酪氨酸磷酸酶受体 Z1 型 ( PTPRZ1 ) 是放射状胶质细胞分化的 GBM 细胞的常见标记基因,基于其在 MES-AC-hybrid 状态下的高速度相图(图 8 G 和 8H,底部和8 I),作者的数据证实了 OPC 和 NPC 样分支内的双向命运。
相关讨论
在过去几年中,通过单细胞分析技术进步取得的众多发现彻底改变了对肿瘤异质性和可塑性以及潜在治疗选择的理解。然而,尚未研究脑肿瘤空间组织的重要性。在这里,作者揭示了微环境与GBM转录异质性的时空变化之间的双向和单向相互作用的见解。揭示了五个空间上不同的转录程序的存在,这些转录程序以共享的转录特征和专有的基因组改变为标志。尽管在 GBM 中有 CNA 的先前描述,但以前没有报道过区域模式和与转录程序的关联。并对对细胞状态、组织学模式和空间专有转录程序进行整体比较。
这种以空间分辨方式在各种分子水平上全面表征 GBM 有助于发现细胞状态的动态适应及其在肿瘤微环境中的空间关系。作者数据表明,在 S 期停止后,严重缺氧等代谢改变会导致增殖受损,从而导致 CNA 的显著积累。这种现象已在其他癌症类型中报道,并且存在足够的证据详细说明缺氧条件下基因组不稳定性的增加。作者在 2D 细胞培养和 TCGA 数据中的结果和验证表明,区域缺氧代谢代表了基因组不稳定性的潜在储存库,并代表了微进化的驱动因素,从而使耐药表型的进化成为可能。
总之,作者阐明了 GBM 的区域转录程序谱,并绘制了它们的微环境景观,包括代谢和肿瘤-宿主细胞相互作用。通过证明宿主环境在重塑遗传和转录异质性方面发挥着重要作用,提供了对患者间异质性的洞察,因此,对于早期 GBM 复发和治疗抗性,需要量身定制的治疗方法,并更加强调个性化方法在神经肿瘤学中的重要性。
小鹿推荐
该团队的工作揭示了GBM中五个空间上组织成分在空间转录组学、空间代谢组学和蛋白质组上不同的特征,证明译患者之间区域共享的转录程序,推断胶质母细胞瘤是由谱系状态的空间分离组织而成的,并可适应炎症或代谢刺激。通过证明宿主环境在重塑转录程序的作用,提供了对患者间异质性、早期GBM复发和治疗的新见解。
在实验技术上方面,文章采用了诸多先进的实验技术,空间转录组(stRNA-seq),空间代谢组学和蛋白组学(成像质谱流式,IMS),这为科研工作者提供了更为广阔的技术视野。在大数据分析方面,作者尝试了很多创新算法,基于机器学习的像素级分类器解决空间精度问题,基于CNA 管道解决拷贝数变异定量问题,这为组学大数据分析领域提供了更多新的见解。在机制探索上,作者联合利用预测算法和体内外模型结合的方式,逐步验证潜在机制,为读者在探索GBM机制方面提供了新的科研视角。
猜你还想看
1、Nature子刊 | 多组学联合分析鼠李糖乳杆菌与白色念珠菌之间的代谢争夺战
2、Environ Int | 浙江大学公共卫生学院周舟课题组在环境科学顶刊首次发文揭示镉暴露与糖尿病的关联
3、项目文章 | LC-MS、GC-MS非靶向代谢组学联合DIA蛋白质组学探究甲状腺毒症小鼠的衰老机制
4、项目文章 | 四川农业大学付凤玲/于好强团队运用TMT磷酸化蛋白质组学探究玉米调控耐旱性机制
END
Q|撰文
小久|排版
欢迎转发到朋友圈
本文系鹿明生物原创
转载请注明本文转自鹿明生物

我知道你在看哟


点击“阅读原文”了解更多
03-08 美谷分子仪器
实验室仪器您做主——免费巡检惊喜来袭!03-08 美谷分子仪器
【中奖通知】元宵节猜灯谜活动中奖名单公示03-08 美谷分子仪器
你真的了解你的护肤品吗?化妆品知识竞答赢女神节好礼!03-08 美谷分子仪器
【精彩回放】人类发育和疾病中的 DNA 甲基化03-08 美谷分子仪器
【产品设置】如何在 SpectraMax i3x 进行光谱扫描检测法的设置?03-08 美谷分子仪器
探索不止,共享卓越 | Opentrons 邀您参加 CACLP 行业盛会03-08 Opentrons
Opentrons 华东区销售经理热力招募,亟待有识之士,共享行业先机!03-08 Opentrons
邀请函 | 高砂电气CACLP2024参展通知03-08 高砂电气
哈克(HAAKE)旋转流变仪在皮肤外用制剂上的应用(文末有惊喜)03-08
仪器租赁 | 赛默飞 气质联用,月租金13220元起03-08
璀璨如你,自成光芒03-08
实验室常见不良习惯揭秘,你中了几个?03-08 实验与分析
【邀请函】东西分析仪器邀您共聚CACLP03-08
活动回顾|东西分析亮相CHINA LAB 2024 展会03-08
3.8国际妇女节 | 致敬每一位“她”03-08
致敬不被定义的她 | 福立仪器祝所有女同胞们节日快乐!03-08 福立仪器
妇女节 | Wǒmen,做自己的样子真美!03-08
短/中链氯化石蜡暴露与妊娠期糖尿病患病风险关系研究03-08
心有光芒,芳华自在,致敬“她力量”03-08