代谢改变是癌症最早观察到的特征之一。最初研究认为,葡萄糖代谢在肿瘤生长中起重要作用,但最近研究发现氨基酸、核苷酸和脂质代谢在癌症中也具有突出重要性。明确限制肿瘤生长的代谢途径为癌症靶向代谢治疗提供依据。研究表明许多癌症丝氨酸合成途径相关酶的表达上调,并且合成途径中第一个酶(磷酸甘油酸脱氢酶,PHGDH)的基因拷贝数增加。然而,丝氨酸合成增加是否促进肿瘤生长以及影响机制存在争议。美国科赫综合癌症研究所Matthew G. Vander Heiden科研团队采用代谢组学等技术研究丝氨酸对肿瘤生长的影响,相关成果发表于《Cell Metabolism》。
与Braf 突变共同促进黑色素瘤形成
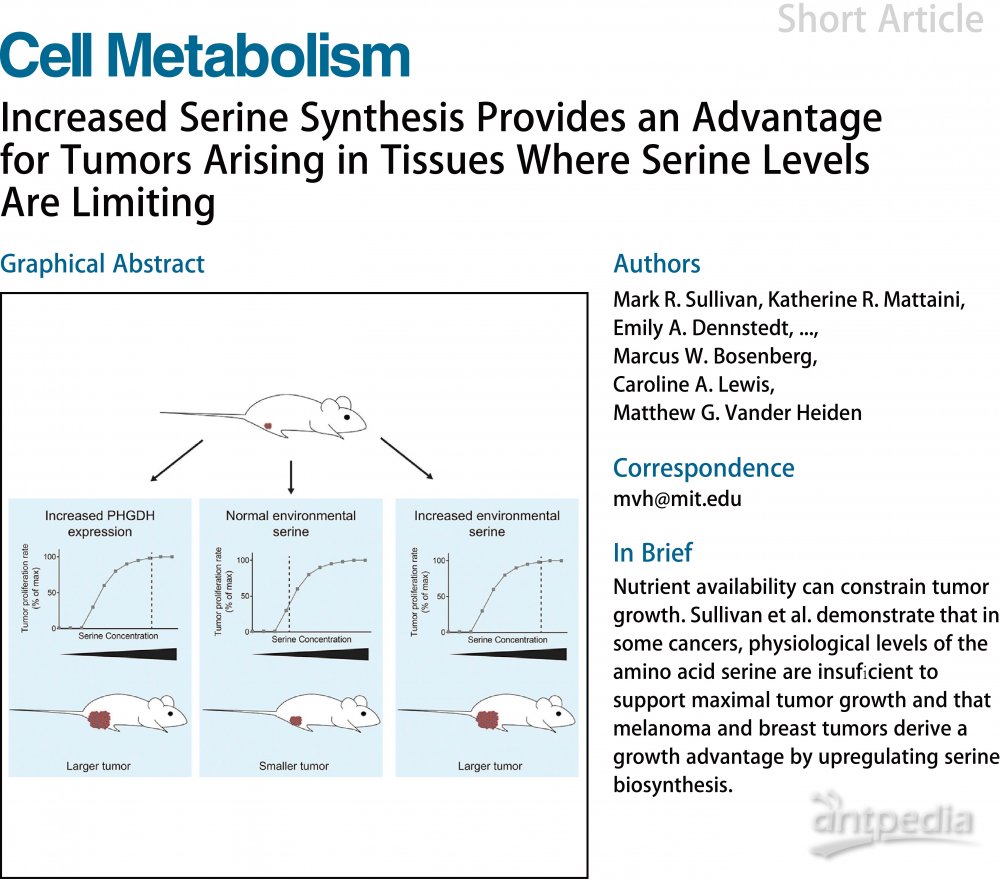
PHGDH在黑色素瘤和三阴性乳腺癌中均表现出基因拷贝数增加,但PHGDH的增加不足以促进肿瘤的发生。因此,PHGDH表达可能与其他遗传事件配合以共同促进肿瘤生长。在黑色素瘤中,研究者发现多达60%的肿瘤中出现Braf 突变。为了确定PHGDH是否可以与Braf 突变体联合促进小鼠黑色素瘤发生,将携带PHGDHtetO等位基因的小鼠与携带BrafV600E条件等位基因(BrafCA)和Tyrl-CreER 等位基因的小鼠杂交并暴露于它莫西芬环境下可使黑色素细胞BrafV600E突变。单独的Braf 活化不会在该模型中驱动肿瘤形成,但当与其他一些遗传改变结合时会促进黑色素瘤形成。当喂食含多西环素的饮食以增加PHGDH表达时,13%的BrafCA; PHGDHtetO ; R26-M2rtTA小鼠形成肿瘤,而不含PHGDHtetO等位基因的黑素细胞Braf活化的同窝小鼠不形成肿瘤(图1A)。肿瘤组织学结果及黑色素瘤标记物Sox10结果(图1B),表明PHGDH表达可与Braf 活化配合共同驱动具有低外显率的黑色素瘤形成。
PHGDH加速黑色素瘤和乳腺癌生长
为了研究PHGDH在黑色素瘤生长和进展中的作用,将PHGDHtetO;R26-M2rtTA小鼠与Braf激活和黑素细胞中的Pten 缺失(BrafCA; Tyr-CreER; Ptenf1/fl)引发的黑色素瘤小鼠杂交。为证实PHGDHtetO等位基因在该模型中表达,采用PHGDHtetO转基因编码人源PHGDH(huPHGDH),其可以与鼠源PHGDH(msPHGDH)区分开。具有PHGDHtetO等位基因的肿瘤将同时表达huPHGDH和内源性msPHGDH,而对照组肿瘤只表达msPHGDH,不表达huPHGDH(图1C)。采用具有相同亲和力的人和小鼠PHGDH的抗体显示PHGDHtetO小鼠肿瘤PHGDH蛋白表达增加(图1D)。 PHGDHtetO等位基因不会改变黑色素瘤的组织学形态(图1E)或血浆丝氨酸水平,但肿瘤达到1cm3 的平均时间从68降低至46天(图1F)。PHGDH表达升高可加速雄性和雌性小鼠中的黑色素瘤生长,其在该模型中表现出不同的肿瘤生长动力学。该表型变化部分原因由细胞增殖增加驱动,因为与对照小鼠相比,具有PHGDHtetO等位基因的肿瘤显示增殖标志物Ki67水平增加(图1G),但未观察到细胞凋亡标记物(剪切型半胱天冬酶3存在差异。总之,PHGDH表达增加可以加速黑色素瘤的生长。
三阴性乳腺癌中也观察到PHGDH拷贝数增加。为了确定PHGDH是否在该癌症中发挥致癌作用,将PHGDHtetO等位基因与Brca1缺失诱导的三阴性乳腺癌Trp53杂合小鼠(BRCAf1/fl;Trp53+/-; MMTV-Cre)杂交。此外,为了证实PHGDH的作用可归因于癌症组织中的PHGDH表达,采用PHGDH高表达仅限于具有乳腺特异性MMTV-rtTA等位基因的乳腺上皮细胞。 PHGDHtetO乳腺癌肿瘤达到1cm3 的时间比对照组快,提示PHGDH表达可促进乳腺癌生长(图1H)。有趣的是,尽管PHGDHtetO小鼠表达人源PHGDH,但末期肿瘤中的PHGDH表达是变化的,且表现出与对照肿瘤相似的PHGDH表达(图1I)。另外,PHGDHtetO肿瘤末期的增殖标志物Ki67或细胞凋亡标记物(剪切型半胱天冬酶3)没有显著差异。一种可能的解释是早期PHGDH表达主要促进早期肿瘤生长。为了证实PHGDHtetO小鼠早期PHGDH表达升高,对7周龄的PHGDHtetO; MMTV-rtTA小鼠和对照MMTV-rtTA小鼠的乳腺进行PHGDH染色。结果发现,与对照小鼠相比,PHGDHtetO小鼠乳房上皮细胞中PHGDH表达增加(图1J)。由此,在肿瘤末期之前,PHGDHtetO等位基因促进肿瘤生长。
乳腺癌细胞系依赖于PHGDH产生丝氨酸
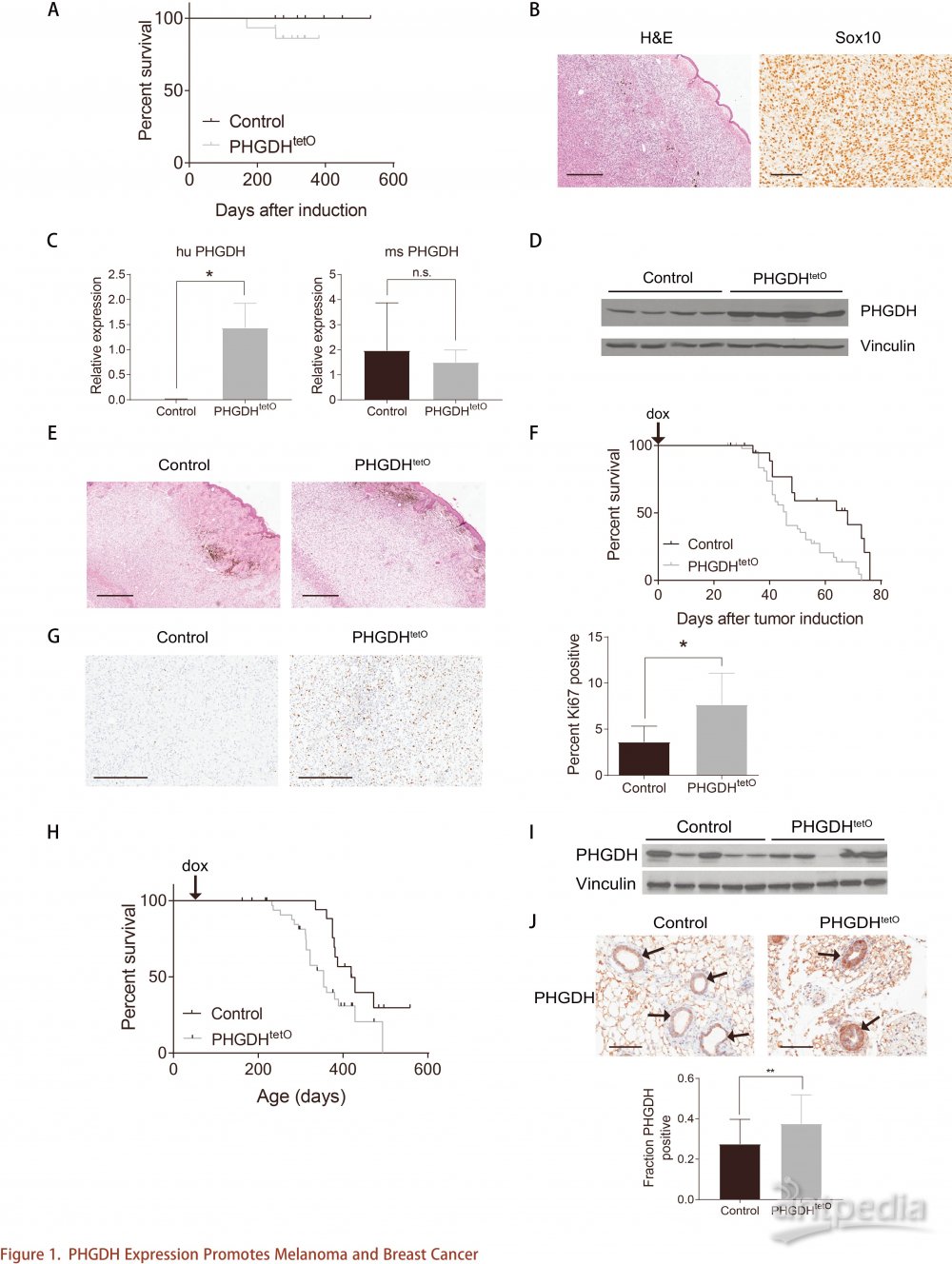
为了确定PHGDH在乳腺癌进程的作用,从对照组末期肿瘤培育细胞系,其PHGDH表达水平与来自PHGDHtetO小鼠的肿瘤相似,并使用CRISPR干扰技术(CRISPRi)敲低PHGDH表达(图2A)。13C 标记的葡萄糖掺入到丝氨酸进行同位素示踪,发现PHGDH敲低后丝氨酸合成途径活性降低(图2B),但对细胞增殖没有影响(图2C)。该结果表明PHGDH可能不是癌细胞增殖所必需的条件。然而,PHGDH表达可能对乳腺癌环境发挥特殊作用,因为,细胞培养和乳腺癌环境之间的一个区别是丝氨酸的可用性。 RPMI-1640培养基中丝氨酸浓度为285mM。相比之下,肿瘤可用的最大丝氨酸浓度可能受血浆中丝氨酸含量的限制,也可能受到营养物质传递中损耗的影响。为了确定在生理条件下丝氨酸的可用性是否限制肿瘤生长,对C57BL/6J 小鼠(C57BL / 6J 小鼠是与本研究中用于原发性黑色素瘤和乳腺癌模型的最相似的鼠种)血浆丝氨酸浓度进行检测。结果显示,血浆丝氨酸水平范围为92至314mM(图2D),类似于人血浆丝氨酸范围(53-262mM)。在生理浓度范围内进行不同浓度丝氨酸培养细胞,观察对照和PHGDH敲低组中的细胞增殖是否有差异,结果显示, PHGDH表达增加仅在较低的丝氨酸浓度下促进细胞增殖(图2E)。由此说明,对于丝氨酸可用性低的组织,丝氨酸的可用性是肿瘤增殖的内源性限制因素。
在喂食小鼠中丝氨酸水平是波动的
小鼠和人体血浆丝氨酸水平的变化表明哺乳动物不能在血液中维持恒定水平,且丝氨酸浓度会随饮食而变化。但是许多哺乳动物,包括老鼠,不会在一天内持续进食。因此,代谢基因表达的昼夜节律效应,以及两餐之间引起的血浆丝氨酸水的波动,都会导致组织中的丝氨酸水平降低,降至一定水平下PHGDH的高表达则促进细胞增殖。为了研究小鼠血浆丝氨酸如何随正常喂养方式而变化,在小鼠自由喂食或禁食情况下,测定C57BL / 6J 和NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ(NSG)小鼠的一天中不同时间点血浆氨基酸水平。结果发现,白天丝氨酸水平达到最高,上午8点和下午5点降至禁食水平(图2F )。其他非必需氨基酸如甘氨酸以及必需氨基酸如赖氨酸也观察到这种模式(图2F)。肿瘤可用的丝氨酸水平会发生波动并周期性接近禁食水平,从而导致丝氨酸在某些情况下受到限制。
饮食可以调节血浆丝氨酸可用性
为了研究体内丝氨酸水平变化是否影响肿瘤生长速度,通过饮食干预调节肿瘤细胞丝氨酸水平。以往研究表明,丝氨酸和甘氨酸缺乏的饮食可以降低血浆丝氨酸水平,从而减缓某些肿瘤类型的生长。由于甘氨酸和丝氨酸可以相互转化,为了确保动物不能从膳食甘氨酸合成丝氨酸,因此饮食中同时去除甘氨酸和丝氨酸。小鼠喂食缺乏丝氨酸和甘氨酸的饮食后,血浆丝氨酸和甘氨酸水平降低并维持了长达30天(图2G)。与对照相比,喂食不含丝氨酸和甘氨酸饮食的小鼠血浆丝氨酸和甘氨酸在一天中也显示出较低水平(图2F)。然而,两者血浆赖氨酸水平相似(图2F),表明氨基酸稳态并非全部发生改变。通过饮食调节可增加丝氨酸水平,喂食丰富丝氨酸饮食的小鼠血浆中具有更高浓度的丝氨酸而甘氨酸没有变化(图2G)。而这些饮食都没有改变小鼠体重(图2H)或在小鼠行为或外观上产生严重异常。并且,不含丝氨酸和甘氨酸的饮食和含有丝氨酸饮食分别稳定保持血浆丝氨酸浓度处于正常生理范围的最小值和最大值范围内。因此,这些饮食可用于研究正常生理范围内的丝氨酸水平对肿瘤生长的影响。
PHGDH提供丝氨酸促进乳腺癌生长
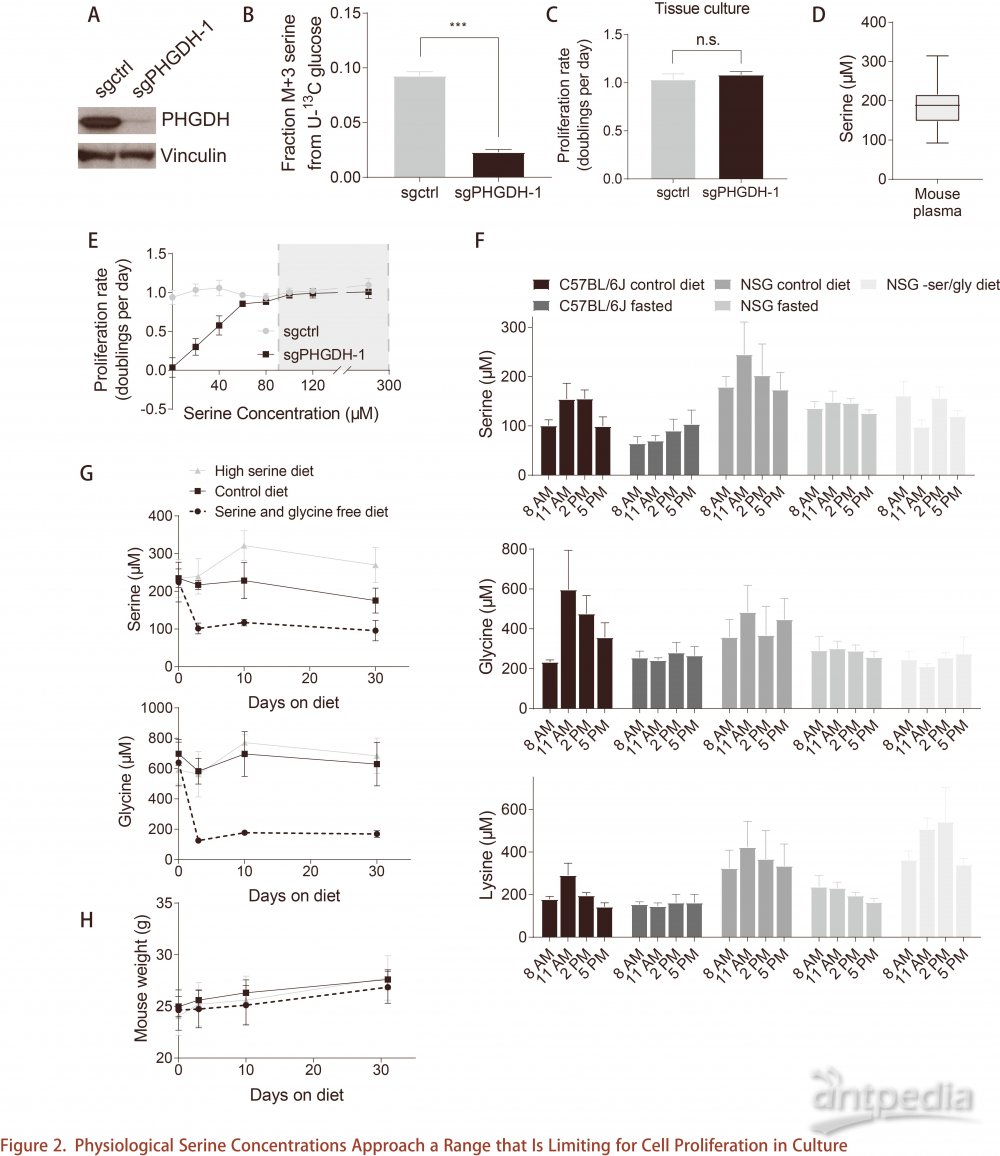
PHGDH能否扭转乳腺微环境中低丝氨酸限制?将源自PHGDH敲低和对照的原发性乳腺癌细胞原位植入NSG小鼠的乳腺脂肪垫。PHGDH敲低的细胞在肿瘤中保留较低的丝氨酸合成途径活性。在正常丝氨酸饮食下,对照细胞组肿瘤比PHGDH敲低细胞组肿瘤生长得更快(图3A),这表明,PHGDH表达可促进乳腺脂肪垫中的肿瘤生长。对照细胞组肿瘤可表达PHGDH,但对丝氨酸饮食不敏感(图3B)。在不含丝氨酸和甘氨酸的饮食下,PHGDH敲低细胞组肿瘤生长更慢(图3C),表明PHGDH低表达的肿瘤对环境丝氨酸水平敏感。富含丝氨酸饮食下的PHGDH敲低细胞组肿瘤与正常饮食下的对照肿瘤生长速度相同,表明丝氨酸饮食能够补偿低PHGDH表达造成的影响。PHGDH敲低细胞组肿瘤中的丝氨酸和甘氨酸水平低于对照肿瘤,而必需氨基酸如缬氨酸和亮氨酸的水平不变,说明PHGDH可以协助肿瘤克服丝氨酸限制(图3D,3E)。这一结果可以解释为PHGDH表达水平发生变化,导致肿瘤中丝氨酸可用性改变,但同时也要考虑这些肿瘤具有不同的增殖率,可能以不同的速率消耗丝氨酸。因此,稳态肿瘤丝氨酸水平不能完全反映丝氨酸的可用性。丝氨酸合成途径也可产生α-酮戊二酸和2-羟基戊二酸,但在PHGDH敲低细胞的肿瘤和对照肿瘤中,两种代谢物水平相似(图3F和3G)。总之, PHGDH活性增加对肿瘤的主要作用是缓冲丝氨酸可用性的波动,使肿瘤细胞免受丝氨酸限制的影响。
不同组织中丝氨酸可用性不同。同时,组织丝氨酸含量包括细胞内和细胞外丝氨酸,不能映出环境中丝氨酸可用性的差异。将来自PHGDH敲低和对照的原发性乳腺癌的细胞植入NSG小鼠的胰腺中,从而进一步研究组织部位的不同是否影响对PHGDH的依赖。胰腺具有比乳腺脂肪垫的丝氨酸水平更高,植入胰腺的对照或PHGDH敲低细胞的小鼠均死于胰腺肿瘤负荷,生存率无显著差异,表明环境丝氨酸水平决定PHGDH表达是否促进肿瘤生长。
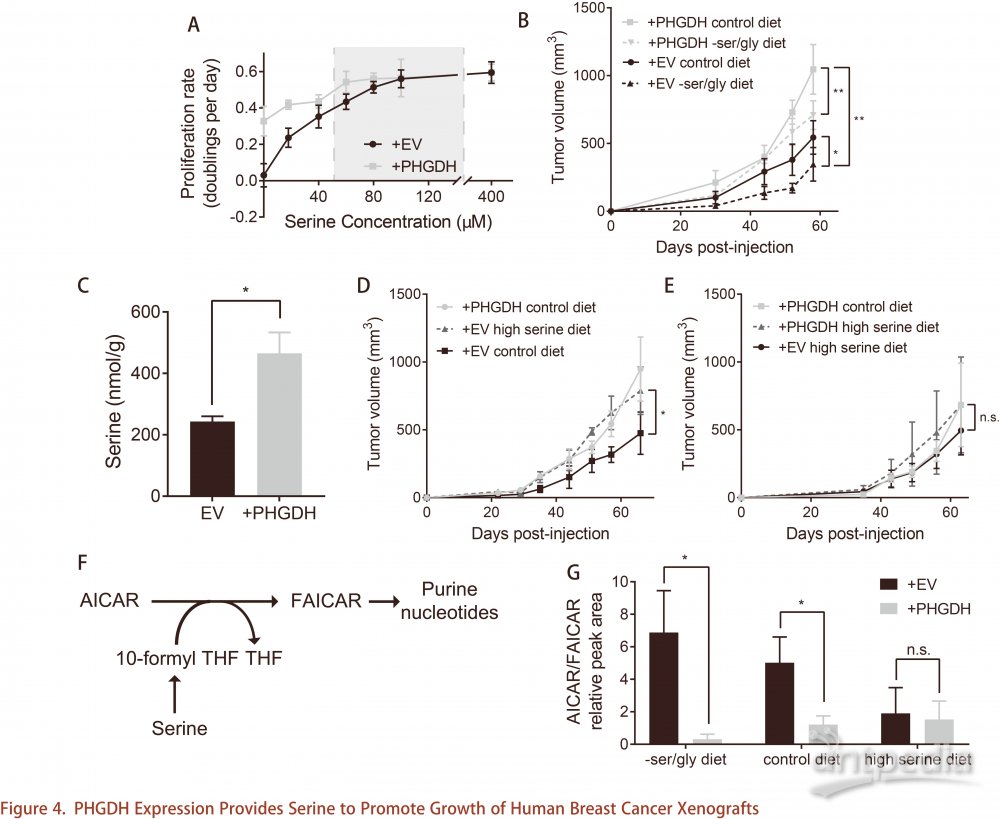
为了确定人类癌症中PHGDH表达增加是否同样提供类似优势,将低表达的PHGDH三阴性乳腺癌细胞MDA-MB-231激活重表达PHGDH,PHGDH表达增加丝氨酸合成途径活性,并且当丝氨酸水平较低时,PHGDH表达为MDA-MB-231细胞提供增殖优势(图4A),与前面结果一致。为了确定这种增殖优势是否在生理环境中也同样存在,将MDA-MB-231细胞原位注射到NSG小鼠的乳腺脂肪垫中。PHGDH敲低细胞形成的肿瘤对丝氨酸和甘氨酸缺乏饮食敏感。此外,缺乏丝氨酸的PHGDH高表达细胞生长速度与喂食丝氨酸饮食的对照细胞生长速度一样快(图4B)。PHGDH表达的肿瘤丝氨酸水平增加,而甘氨酸,α-酮戊二酸或总2-羟基戊二酸水平不变(图4C)。这表明丝氨酸是MDA-MB-231乳腺脂肪垫肿瘤生长的限制因素。丝氨酸饮食小鼠肿瘤细胞比正常饮食小鼠中的肿瘤细胞生长更快(图4D)。而且, PHGDH高表达小鼠正常饮食与对照小鼠丝氨酸饮食相比,两者细胞的生长速率相同,表明PHGDH表达或丝氨酸饮食两者中的任一条件都足以克服丝氨酸限制。此外,无论是丝氨酸饮食小鼠,亦或正常饮食PHGDH高表达小鼠,还是丝氨酸饮食PHGDH高表达小鼠的,三者肿瘤生长速度相同(图4E)。总之,PHGDH和丝氨酸饮食的效果是冗余的, PHGDH通过向低丝氨酸肿瘤环境中的细胞提供丝氨酸来促进肿瘤生长增加。
PHGDH维持丝氨酸依赖性生物合成过程
如果在生理条件下肿瘤生长遇到丝氨酸不足,需要丝氨酸的生物合成过程就会出现扰动。丝氨酸是一碳单位供体,对下游过程至关重要,包括核苷酸合成、脂质合成和SAH再生SAM,这些过程都有助于肿瘤生长。嘌呤核苷酸合成中的多个反应需要一碳单位,例如,由5-氨基咪唑-4-甲酰胺核糖核苷酸(AICAR)生成5-甲酰胺基咪唑-4-甲酰胺核糖核苷酸反应(FAICAR)(图4F)。丝氨酸不足将降低一碳单位并且增加AICAR水平。研究结果显示,对照组中AICAR与FAICAR的比值高于PHGDH高表达组(图4G),这与PHGDH表达的肿瘤更有能力产生FAICAR一致。这种效应在喂食不含丝氨酸和甘氨酸饮食的小鼠和正常饮食小鼠均存在(图4G)。相反,丝氨酸饮食小鼠的AICAR/FAICAR比值不存在PHGDH依赖性,也不存在PHGDH依赖性生长优势。这表明丝氨酸受限的肿瘤可能缺乏一碳单位,从而抑制肿瘤增殖。PHGDH表达或提供丝氨酸饮食都改变AICAR/FAICAR比率,表明PHGDH为生物合成过程提供丝氨酸从而促进肿瘤生长。
小结
癌症中会发生PHGDH表达增加,这样一来,在丝氨酸限制的组织环境中细胞也会加速增殖。PHGDH表达或饮食变化补充丝氨酸的可用性会为黑色素瘤和乳腺癌提供增殖优势。此外,缺乏丝氨酸饮食不会抑制小鼠胰管腺癌生长,并且PHGDH表达不影响植入胰腺的乳腺细胞的肿瘤生长。这表明,不同组织环境中肿瘤生长受到不同营养素可用性的限制。丝氨酸限制是否通过削弱特定生物合成途径活性来降低肿瘤生长尚不清楚。而且,丝氨酸的可用性不是限制肿瘤生长的唯一代谢参数,其他代谢途径也可以影响微环境中较低的营养素水平。研究哪些代谢物和途径限制肿瘤生长可以更好的解释癌症发生的机制,并有利于指导我们采取何种饮食干预措施以抑制癌症的发生发展。
参考文献
Sullivan et al., Increased Serine Synthesis Provides an Advantage for Tumors Arising in Tissues Where Serine Levels Are Limiting. Cell Metab. 2019 . doi: 10.1016/j.cmet.2019.02.015.

