400-6699-117转1000
咨询列表
上海鹿明生物科技有限公司
您好,欢迎您查看分析测试百科网,请问有什么帮助您的?

诚信认证:
工商注册信息已核实! 扫一扫即可访问手机版展台
扫一扫即可访问手机版展台
| 参考报价: | ¥800-1000 RMB(人民币) | 型号: | GCMS非靶向代谢组学分析 |
| 品牌: | 鹿明生物 | 产地: | 上海 |
| 关注度: | 752 | 信息完整度: | |
| 样本: | 典型用户: | 暂无 |
400-6699-117转1000
简介
非靶向代谢组学
非靶向代谢组学又称全代谢组学(Untargeted metabolomics),对样本进行全面的、无侧重的检测,通过比较实验组和对照组样本,找出差异代谢产物,结合生物信息分析,找出富集差异通路,整合代谢网络。全代谢组旨在提供一个全面的研究视角来比较差异,通常用于标志物的筛选、分子机制研究及多组学整合分析。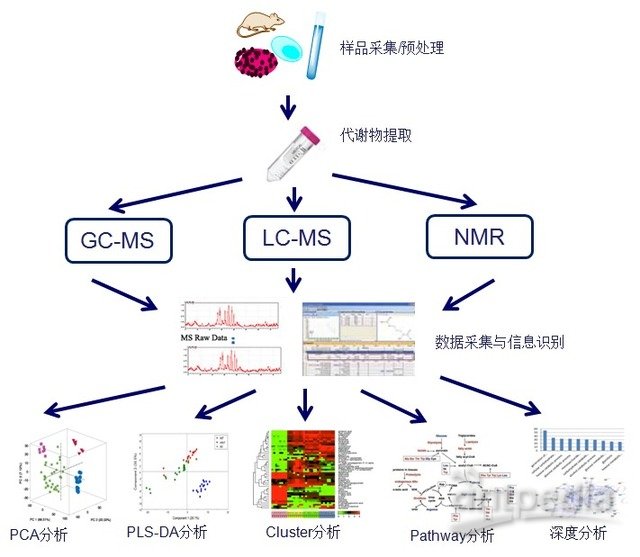
技术特征 非靶向代谢组学
GC-MS(气相色谱)
GC-MS是代谢组学研究的经典技术,具有技术成熟稳定、灵敏度高等特点,同时,由于具有相对完善的数据库,使得定性也更为准确可靠,缺点则主要表现在样品处理相对复杂,对于不易衍生化的物质定性和定量较困难。
LC-MS(液相色谱)
优势主要表现在样品制备和前处理简单、实验重复性好,灵敏度高,分离和分析范围广,缺点则主要表现在数据库完善程度不够,定性相对困难。
NMR(核磁共振)
最大的优势是对样品无破坏性,测定无偏向性,即适用于血液、尿液体液等液体样品,也适用于组织和器官等固体样品,并且测定速度快,可实现样品代谢组的动态监测,缺点是灵敏度低。
技术优点 非靶向代谢组学 
实验流程 非靶向代谢组学: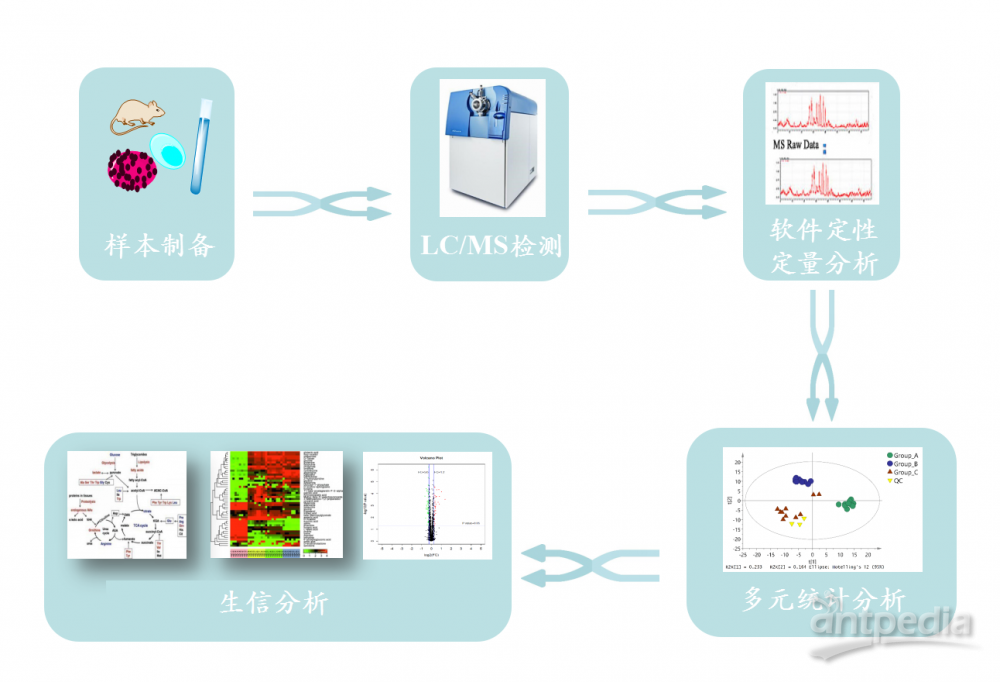
分析内容: 非靶向代谢组学
送样要求: 非靶向代谢组学 
仪器设备: 非靶向代谢组学: 
数据分析 非靶向代谢组学
PCA(主成分)统计分析
主成分分析是代谢组数据常用的分析方法之一。通过主成分分析,可以了解不同代谢产物在区分不同样品中的权重,对于差异代谢产物的筛选特别是重要分子标志物的筛选具有极为重要的意义和价值。
PLS-DA(偏最小二乘法-判别分析法)统计分析
偏最小二乘法- 判别分析法(PLS-DA)是模式识别的重要方法。该方法能够从复杂的数据中最大限度地提取简化后的信息以供分析。
Cluster-heatmap聚类分析
聚类分析旨在利用数据分布的相似性进行分类,是模式识别的重要方法。该方法能够从复杂的数据中最大限度地提取简化后的信息以供分析。
代谢物功能分析
在差异代谢产物筛选的基础之上,分析代谢产物的生物学意义是代谢组学研究的最终目的。我们可以通过PCA、聚类等统计学分析结合Pathway回溯,代谢产物和蛋白/基因互作等生物信息学分析对代谢产物在生物体内的功能和作用有更深入的了解。
GC-MS非靶向代谢组学客户文章实例 GC-MS非靶向代谢组学
前言
农业增产及产品品质提升一直都是生物课题研究中重要的一部分,特别是稀有和低产物种。白灵菇是一种珍贵的珍稀食用菌,具有极高的营养和药用价值。本文为鹿明生物合作客户中国农业科学院农业资源与农业区划研究所胡清秀博士团队在Frontiers in Microbiology发表的 “白灵菇菌丝体生理成熟过程中的代谢轮廓谱分析及潜在菌丝体指示因子的探索”的文章。文章报道通过GC-MS非靶标代谢分析,发现N-氨基甲酰基L-天冬氨酸(CA-asp)可作为评估菌丝体成熟度的潜在指标,揭示了白灵菇菌丝体生理成熟期的代谢调控网络变化机制。

英文标题:Metabolic Profiling of Pleurotus tuoliensis During Mycelium Physiological Maturation and Exploration on a Potential Indicator of Mycelial Maturation
中文标题:白灵菇菌丝体生理成熟过程中的代谢轮廓谱分析及潜在菌丝体指示因子的探索
材料:白灵菇
影响因子:4.259
发表期刊:Frontiers in Microbiology
主要运用鹿明生物技术:GC-MS非靶标代谢组
研究背景
白灵菇是一种珍贵稀有的食用菌,具有极高的营养和药用价值。由于栽培技术尚不成熟,导致产量和质量波动较大。培育难点在于评估菌丝体的成熟度,目前缺乏可明确表征菌丝体生理成熟程度的量化指标。本研究的目的是通过GC-MS代谢组学的方法来研究白灵菇菌丝体生理成熟的潜在指标,进而改进白灵菇栽培方法,最大限度地提高产量和质量。
研究思路

研究方法
1. 实验材料培育及分组
预先培养的白灵菇菌丝体接种,并在培养瓶中以25℃(黑暗中)的条件培养25天,之后分为两组,分别在17℃和29℃的温度下度过菌丝体生理成熟期(MPMP,35天)。此后将所有瓶子转移到蘑菇屋(温度10-15℃,相对湿度90–95%)培养,以诱导原基分化和子实体发育,记录包括伞盖直径、结实率、总产量和生物效率指标。在MPMP之前和MPMP第35天(17℃和29℃)采集菌丝体(表1)并进行代谢组学分析,每个时间点8个生物学重复。
表1 |菌丝体采样信息

2.检测方法
(1)GC-MS非靶标代谢组学技术
(2)菌丝体中N-氨基甲酰基L-天冬氨酸(CA-asp)含量的测定:PDAB(对二甲甲醛)显色法
实验结果
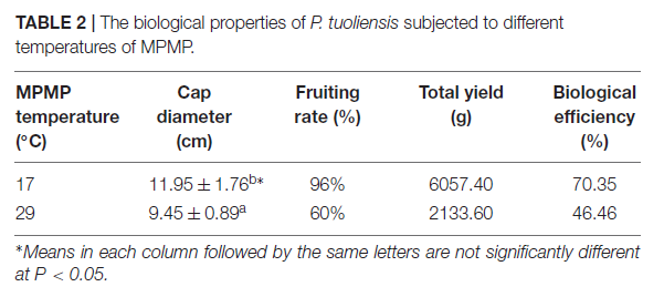
1.不同MPMP温度对白灵菇生物学特性的影响
记录菌盖直径、结实率、总产量和生物效率,以评估不同MPMP温度(17和29℃)对白灵菇发育的影响(表2)。伞盖直径是衡量子实体质量的重要指标,直径越大,质量越高。如图1所示,在17℃的子实体的伞盖直径明显大于29℃,在29℃下的结实率和总产量显著低于17℃,同样在17℃下的生物效率比在29℃高。

图1 | 分别在(A)17 ℃和(B)29 ℃下菌丝体生理成熟期 35天后的子实体直径
表2 |不同菌丝体生理成熟期温度下的白灵菇生物学特性
2.基于GC-MS的生理成熟菌丝体的代谢特征
为了调查MPMP过程中菌丝体的代谢变化,采用GC-MS代谢组学方法分析了第0天、第35天17℃和第35天29℃菌丝体的细胞外液样品(A、B和C样品)。三个样品共鉴定出236种代谢物。采用多元统计分析的方法,处理数据并得到差异代谢物。
3.多元统计分析
首先采用无监督的PCA分析来评估所有样品之间的总体分布以及整个分析过程的稳定性。如图2所示,PCA图的四个主要成分解释了59.1%的总方差信息。所有的QC(质量控制)样品均聚在一起,表明具有良好的分析稳定性和实验可重复性。在PCA模型中,三组样品在第一组分之间很好地分离,表明三组之间具有明显的代谢多样性。其中菌丝体A样品浓度更高,与菌丝体B和菌丝体C样品略有不同。这些结果与先前推测一致,即MPMP前后菌丝体细胞外液中的代谢谱差异很大。
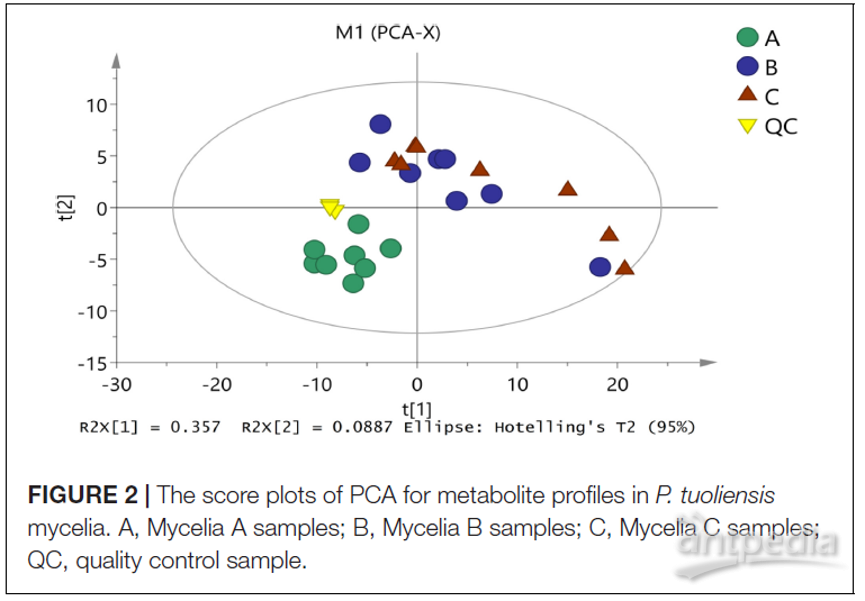
图2 | PCA得分图显示了白灵菇菌丝体中的代谢轮廓谱
A,菌丝体A;B,菌丝体B;C,菌丝体C;QC,质量控制样品
有监督PLS-DA模型显示,菌丝体A,B和C样品之间存在明显代谢差异(图3A,C,E)。PLS-DA模型的拟合度和预测能力均比较可信(表3),并未过拟合(图3B,D,F)。
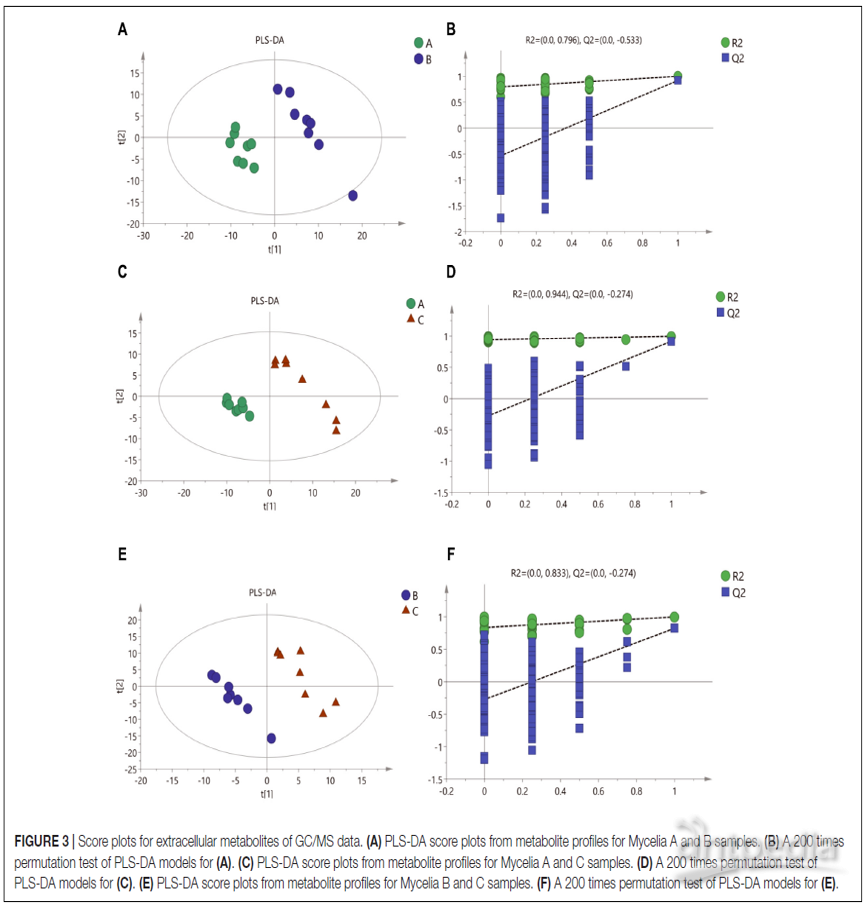
图3 | 基于GC / MS数据的细胞外代谢物得分图
(A)来自菌丝体A和B样品代谢轮廓谱的PLS-DA得分图;
(B)对(A)的PLS-DA模型进行200次排列检验;
(C)来自菌丝体A和C样品代谢轮廓谱的PLS-DA评分图;
(D)对(C)的PLS-DA模型进行200次排列检验;
(E)来自菌丝体B和C样品的代谢轮廓谱的PLS-DA得分图;
(F)对(E)的PLS-DA模型进行200次排列测试。
表3 |PCA模型参数
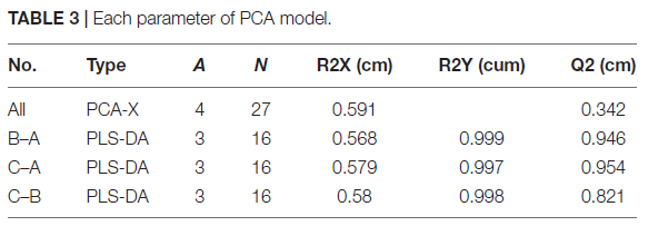
4.差异代谢物
使用VIP值对样品之间的差异代谢物进行筛选,并通过非参数Mann-Whitney U检验进行确认。在三组样品中共鉴定出117种VIP值大于1.0且p <0.05的差异代谢物,包括41种有机酸、21种氨基酸、14种醇、11种糖、8种胺、5种酮、1种酯和16种其他代谢物。在MPMP第0天收集的菌丝体中很少检测到某些代谢物(A样品),而在第35天收集的菌丝体(B和C样品)中却显示出大量增加的代谢物,例如CA-asp、柠康酸、二氢香芹酚、褪黑激素和奎丁-4-羧酸。根据VIP值大于1.0和p <0.01的标准鉴定出72种差异代谢物(表4)。
表4 通过GC-MS鉴定的菌丝体A,B和C样品之间的差异代谢物
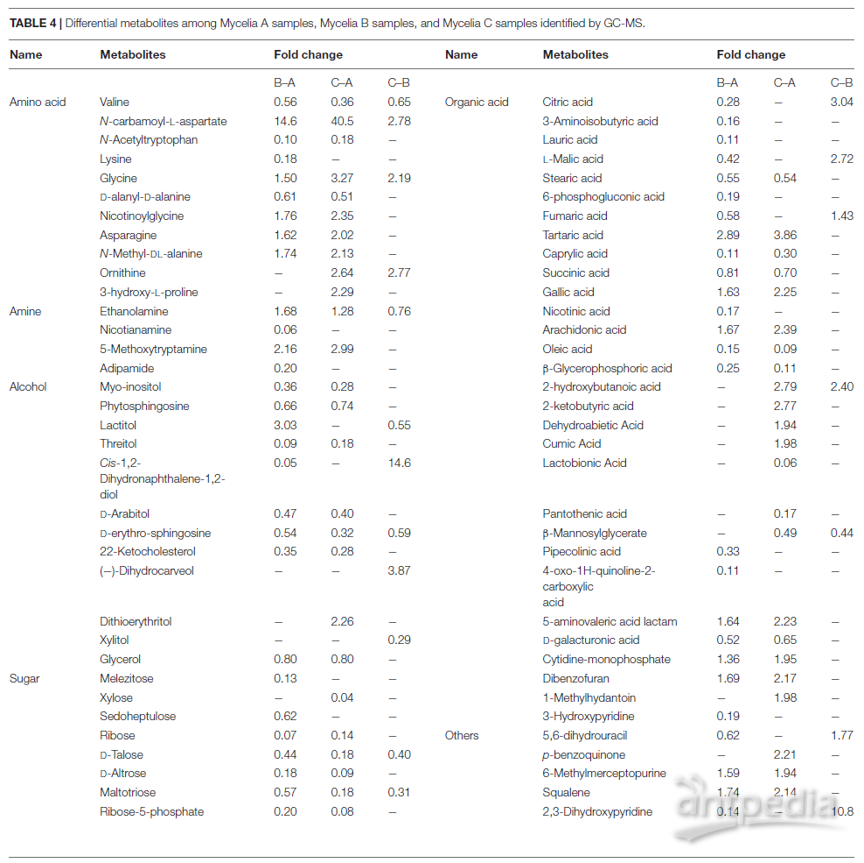
在菌丝体A,B和C样品的热图中观察到了差异代谢物明显的动态变化(图4A)。图4A的上部显示了与菌丝体A样品相比,菌丝体B和菌丝体C样品中丰度明显降低的代谢物,而图4A的下部显示了与菌丝体A样品,例如CA-asp、甘氨酸、乳糖醇、酒石酸、内酰胺、角鲨烯、6-甲基巯基嘌呤、5-甲氧基色胺、天冬酰胺、没食子酸和烟酰甘氨酸。图4B显示了与菌丝体B样品相比菌丝体C样品中的差异代谢产物。与菌丝体C样品相比,菌丝体B样品中丰度显著增加的代谢物包括缬氨酸、6-磷酸葡萄糖酸、D-塔洛糖、β-甘露糖基甘油酸酯、木糖醇、半乳糖醇、乳糖醇和麦芽三糖。而(-)-二氢香芹酚、2-羟基、柠檬酸、5,6-二氢尿嘧啶、L-苹果酸、马来酸、甘氨酸和鸟氨酸的丰度显著降低。
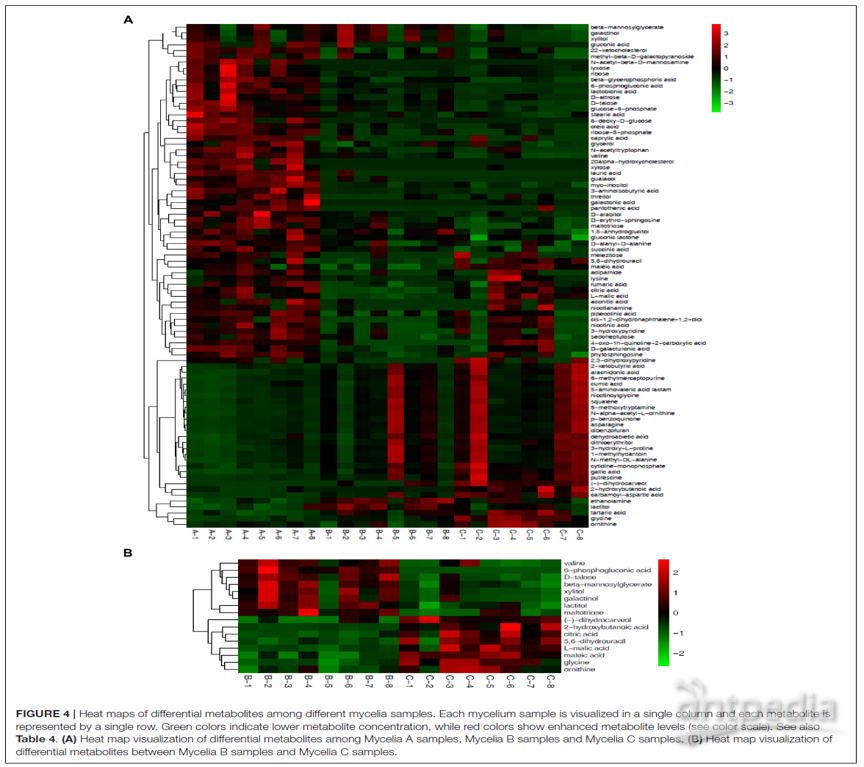
图4 | 不同菌丝体样品之间差异代谢物的热图
(A)菌丝体A,B和C样品之间差异代谢物的热图;
(B)菌丝体B样品和菌丝体C样品之间差异代谢物的热图;
列代表每个菌丝体样品,行代表每个代谢物;绿色表示较低的代谢物浓度,而红色表示较高的代谢物水平,参见表4。
图5显示了菌丝体B样品和菌丝体A样品之间(A),菌丝体C样品和菌丝体A样品之间(B)以及菌丝体C样品和B样品之间(C)的代谢物的相关性分析。较大的值表明两种代谢物之间的相关性更大。

图5 | 差异代谢产物之间的相关性分析图
(A)菌丝体A和B样品,(B)菌丝体C和A样品,(C)菌丝体C和B样品。
蓝色代表正相关,而红色代表负相关,值越大表明两种代谢物之间的相关性更大。
5.代谢通路分析
为了深入了解三组样品之间代谢网络的差异,将确定的72种差异代谢物进行代谢途径富集分析。图6显示了菌丝体B和菌丝体A样品,菌丝体C和菌丝体A样品以及菌丝体C和菌丝体B样品之间的富集途径,包括五个主要的代谢途径——糖酵解、有机酸代谢、氨基酸代谢、三羧酸循环(TCA)和糖代谢。以及一些次级代谢途径——烟酸和烟酰胺代谢、氧化磷酸化。
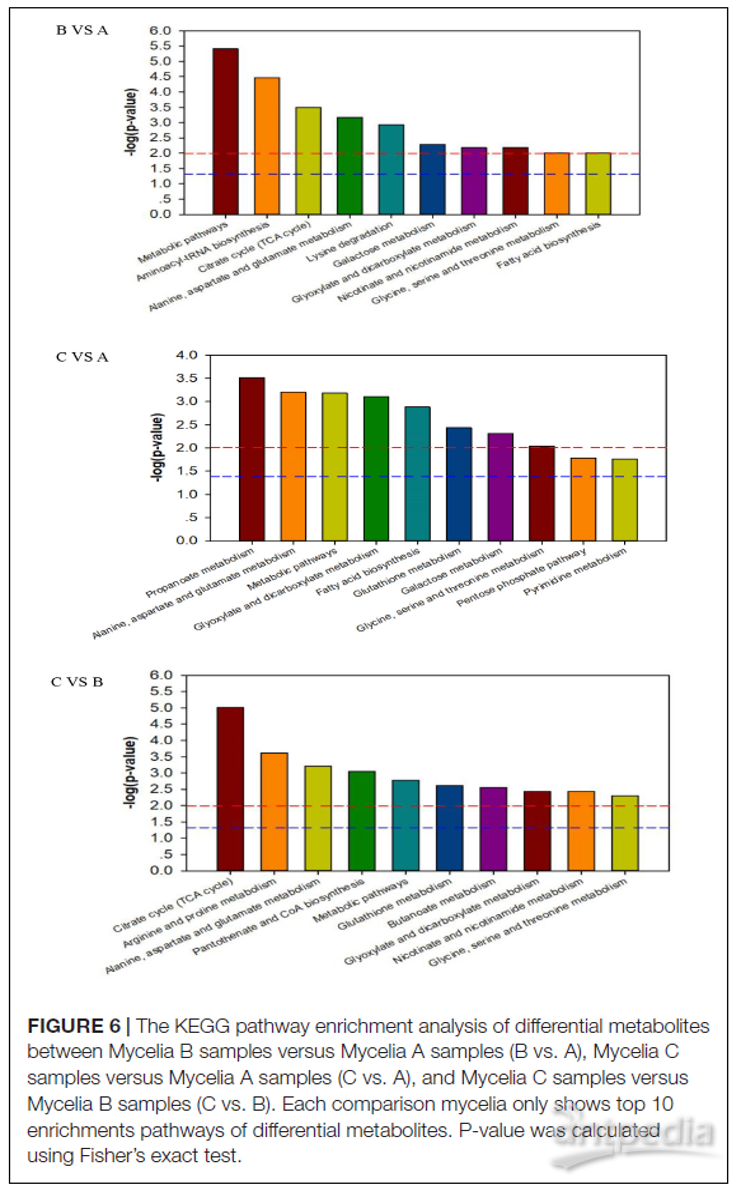
图6 | 差异代谢物的KEGG途径富集分析图
B vs. A菌丝体B与A样品,C vs. A菌丝体C与A样品,C vs. B菌丝体C与B样品。每组比较的菌丝体仅显示前10个差异代谢物的富集途径。P值使用Fisher精确检验计算得出的
图7显示了每种代谢途径中涉及的一些显著改变的途径和差异代谢物。
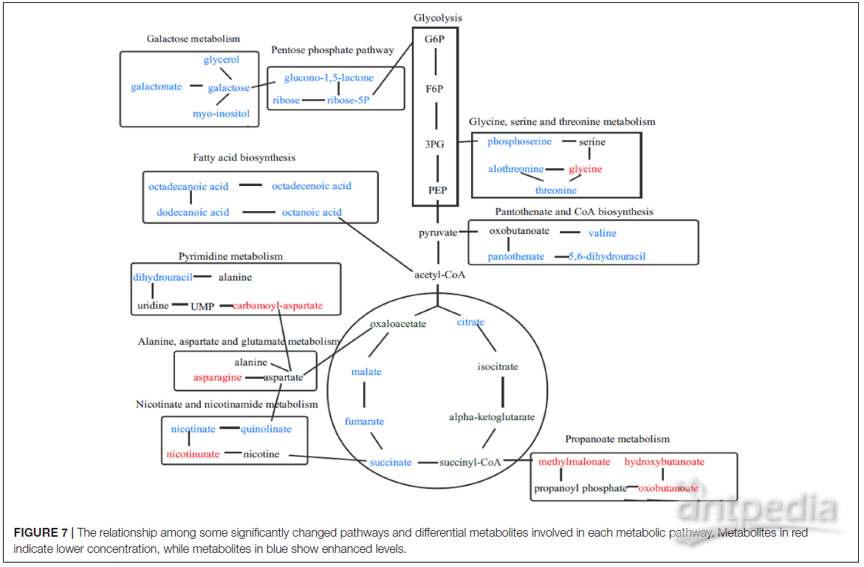
图7 | 某些显著改变的途径与每条代谢物途径所涉及的差异代谢物之间的关系
红色表示代谢物浓度较低,而蓝色表示代谢物浓度升高
图8仅显示了在MPMP第35天被显著激活的整个嘧啶代谢途径。
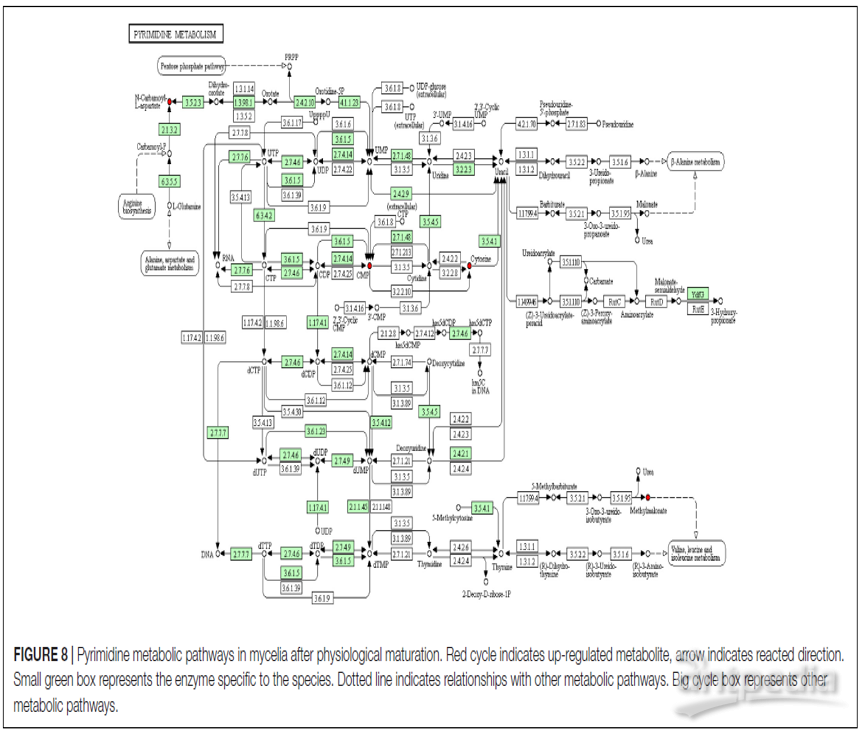
图8 | 生理成熟后菌丝体的嘧啶代谢途径
红色循环表示代谢物上调,箭头表示反应方向,小绿框代表该物种特有的酶,虚线表示与其他代谢途径的关系,大循环框代表其他代谢途径
6.在不同MPMP阶段收集的菌丝体样品中CA-asp含量
为了评估CA-asp作为菌丝体生理成熟指标的可行性,在不同MPMP阶段收集的菌丝中测量CA-asp含量。如图9所示,在MPMP过程中CA-asp含量逐渐增加。刚进入生理成熟期(MPMP第0天)时,其含量仅为0.21 g / L。而在MPMP第45天时其含量上升至1.55 g / L,此后缓慢上升并在1.60 g / L时升高。MPMP第50天,但与第45天的CA-asp含量相比,后者的变化并不明显。
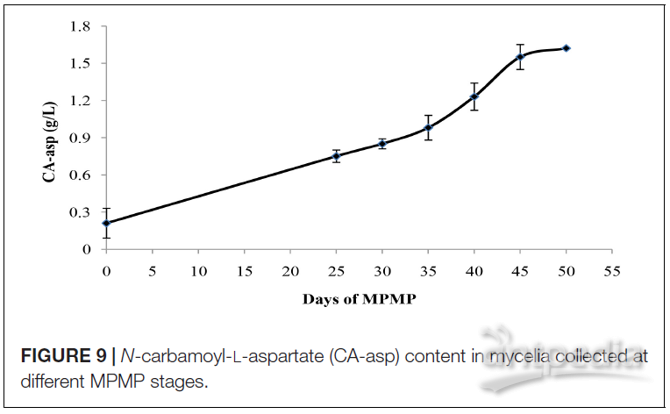
图9 | 不同的菌丝体生理成熟期收集的菌丝体中N-氨基甲酰基-L-天冬氨酸(CA-asp)含量
相关讨论
本实验发现在17℃持续35天的MPMP可导致伞盖直径更大、结实率更高、产量增加和生物效率更高,但是较低温度促进子实体发育的原因尚不清楚。在MPMP期间,菌丝体中的某些代谢物发生很大变化。因此它们可能是进入原基分化阶段的重要信号。代谢组学为研究MPMP前后以及温度胁迫期间菌丝体的代谢谱提供了一种有效的方法。作者进行了基于GC-MS的非靶标代谢组学研究,考察了MPMP第0天、第35天(17℃和29℃)菌丝体中代谢产物的变化,以寻找菌丝成熟的潜在生物标记物。研究结果表明代谢物曲线受菌丝体生长以及温度胁迫的影响,三组菌丝体样品之间的72种差异代谢产物发生了很大变化(p <0.01和VIP> 1.0)。代谢途径富集分析的结果表明,许多鉴定出的代谢物参与糖代谢,氨基酸代谢和三羧酸(TCA)循环。生理成熟的菌丝体中柠檬酸、根琥珀、富马酸、苹果酸根和核糖的减少表明,TCA周期降低,菌丝体代谢减慢。因此作者推测减慢新陈代谢是为了防止菌丝体过度分解,从而避免在原基和子实体形成过程中营养不足。
成熟菌丝体中糖的浓度降低,此现象与真菌菌丝体的生长情况一致。担子菌,例如牡蛎蘑菇、冬菇和香菇,可以将含有木质素,半纤维素和纤维素的栽培底物酶解为低分子量的糖,然后被真菌的菌丝吸收以支持其生长和成熟,这一过程称为营养吸收。在菌丝成熟的后期,新陈代谢的减慢导致细胞外酶水平的降低,因此降解产物的丰度相应降低。本研究中在成熟菌丝体中观察到极低浓度的核糖(戊糖磷酸途径的关键中间体),表明该途径也被抑制。磷酸戊糖途径是一种生物化学途径,与糖酵解同时起作用,并产生烟酰胺腺嘌呤二核苷酸磷酸(NADPH)和戊糖(5-碳糖),并涉及葡萄糖的氧化。因此它的主要作用是合成代谢而不是分解代谢。磷酸戊糖途径是生物合成过程(如脂肪酸合成和无机氮同化)还原剂的主要来源。在本研究中,脂肪酸合成途径中辛酸、十二烷酸、十八烷酸和十八烯酸的丰度在MPMP第35天急剧下降。用于脂肪酸合成的还原剂主要来自戊糖磷酸途径,这些脂肪酸的下调与磷酸戊糖途径中间体的下调高度吻合。
在该研究中鉴定出的大多数氨基酸在MPMP第35天的菌丝体中含量明显增加,这表明氨基酸代谢在菌丝体成熟过程中至关重要。在MPMP第35天时,可以通过糖异生作用转化为葡萄糖的糖原氨基酸(甘氨酸)的丰度显著提高(p <0.05)。天冬氨酸主要来自草酰乙酸,菌丝体成熟后天门冬氨酸的含量变化不明,但是天冬氨酸的前体——天冬酰胺和CA-asp在成熟菌丝体中大量积累,天冬酰胺和CA-asp丰度的增加归因于其他代谢途径的积极参与,而不是因为涉及天冬氨酸的丙氨酸,天冬氨酸和谷氨酸代谢的存在。
作者观察发现,与CA-asp相关的嘧啶合成被激活,这表明菌丝体的生理成熟增加了对核苷酸的需求,用来快速合成细胞材料,这是通过从头合成和/或核苷酸拯救来实现的。图8显示了生理成熟后菌丝体的整个嘧啶代谢途径。成熟的菌丝体显示核苷酸浓度升高,包括胞苷一磷酸(CMP)和胞嘧啶,这表明在生理成熟过程中对快速核苷酸合成的需求很高。丙二酸甲酯是酸甲酯催化的降解产物,与缬氨酸,亮氨酸和异亮氨酸代谢有关。成熟菌丝体中丙二酸甲酯的丰度增加可能意味着相应的代谢网络比未成熟菌丝中的代谢网络更活跃。此外细胞外核苷酸通过嘌呤能信号传导刺激菌丝体存活。
如上所述,MPMP温度会影响子实体的产量和质量,这也反映在代谢产物丰度的差异上。在MPMP第35天,菌丝体C样品(29℃)中柠檬酸、马来酸和L-苹果酸的浓度显着高于菌丝体B样品(17℃)的浓度(图4B)。这些不同的代谢物参与了TCA循环,与能量代谢有关。因此菌丝体C样品的代谢可能比菌丝体C样品更活跃,这表明菌丝体C样品消耗了更多的培养底物,从而导致营养成分浓度不足,无法形成原基和子实体。这种差异也解释了为什么在生理成熟过程中暴露于低温下的菌丝体伞盖直径更大、结实率更高、产量增加和生物学效率更高。
在鉴定出的代谢产物中,作者极其关注CA-asp。因为其菌丝体B和菌丝体C样品的丰度与其菌丝体A样品的丰度相比存在高倍变化差异。有趣的是,在MPMP的第0天收集的菌丝体中几乎未检测到CA-asp,但在生理成熟的菌丝体中含量极高。因此作者推论CA-asp可以用作鉴定菌丝体是否达到生理成熟的潜在标记物。作者还检测了不同MPMP阶段菌丝体中CA-asp的含量,发现CA-asp含量持续上升直至MPMP的第50天,菌丝体中CA-asp的含量在生理成熟过程中显示出规律变化。
实验结论
白灵菇的生物特征结果表明较低的MPMP温度(17℃)对其生长发育有利。采用基于GC-MS的非靶标代谢方法来比较在MPMP第0天、第35天(17℃和29℃)菌丝体中代谢产物的丰度,鉴定出236种差异代谢产物,根据VIP大于1和p <0.01的标准选择出72种差异代谢产物(上调37.8%,下调62.2%)。这些代谢物主要参与糖酵解、有机酸代谢、氨基酸代谢、TCA循环、糖代谢、烟酸酯和烟酰胺代谢以及氧化磷酸化。嘧啶代谢途径可能与菌丝体的生理成熟有关,参与该途径的代谢产物CA-asp的含量在MPMP第35天显著增强,是菌丝体成熟的潜在指标。作者证明了CA-asp在不同MPMP阶段收集菌丝体中的含量呈规律性变化,这表明CA-asp可用作菌丝体成熟指标的可行性,但该指标是否可用于商业生产需经大规模验证。
小鹿推荐
本文运用基于GC-MS的非靶标代谢组学技术发现潜在生物标志物,进行植物生长规律的研究。研究过程思路清晰、数据详实,结合生理指标参数,找到了可以反映菌丝体成熟度的特征代谢物,是一篇典型的非靶标代谢组学文章。
小鹿智囊 非靶向代谢组学
1、代谢组研究相对于基因组和蛋白质组研究而言有什么不同之处?
主要特点包括三方面:一、基因和蛋白表达的有效的微小变化会在代谢物上得到放大,从而使检测更容易;二、代谢组学的技术不需建立全基因组测序及大量表达序列数据库;三、因为代谢产物在各个生物体系中都是类似的,所以代谢组学研究中采用的技术更通用。
2、代谢组的样品重复性有什么要求?
代谢组学基于多元统计分析方法进行的,在样品准备上相对于转录组和蛋白质组而言需要更多的重复数据。一般建议植物样品:最少6次,建议8次生物学重复;
模式动物及微生物样品:最少8次,建议10次生物学重复;
临床样品:30次生物学重复以上,如组织样品不好取样,可控制在20次重复以上。
3、靶向代谢检测平台的选择?
GC-MS和LC-MS两种平台各有优势,主要是由目标化合物的理化性质决定,GC-MS适合易挥发性、分子量不太大的化合物,而且,离子源为EI,碎片信息多,有成熟的图谱库,便于化合物检索,对于非挥发性成分则需要衍生化;而LC-MS离子源为APCI或者ESI源,适合做中等或强极性的化合物,分子量范围广,灵敏度高,前处理简单,但是LC-MS的价格要贵点,因此可以根据实际情况选择合适的检测平台。
4、关于代谢组数据库问题,老师经常问起数据库是商业库还是自己建的库,类型?
GC-MS数据库:Nist、feihn;
LC-MS数据库:HMDB、Metlin、自建库(大连化物所-许国旺组)。
GCMS非靶向代谢组学分析信息由上海鹿明生物科技有限公司为您提供,如您想了解更多关于GCMS非靶向代谢组学分析报价、型号、参数等信息,欢迎来电或留言咨询。
注:该产品未在中华人民共和国食品药品监督管理部门申请医疗器械注册和备案,不可用于临床诊断或治疗等相关用途